Stegosauridae
Stegosauridae è un clado estinto di dinosauri tireofori facenti parte del sottordine di Stegosauria. Il clado è definito come "tutte le specie di dinosauri più strettamente legate a Stegosaurus che a Huayangosaurus".[6] Il nome "stegosauridae" è quindi un nome staminale che prende il nome dal suo genere meglio rappresentato, ossia Stegosaurus (il cui nome significa "lucertola tetto").[7] Sulla base delle prove fossili, gli stegosauridi vissero dal Giurassico inferiore fino al Cretacico inferiore , in quelli che oggi sono il Nord America, Eurasia e Africa. D'altra parte, il sister taxon di stegosauridae, gli Huayangosauridae vissero solo durante il Giurassico medio.[8][9]
| Stegosauridae | |
|---|---|
 | |
| Stato di conservazione | |
Fossile | |
| Classificazione scientifica | |
| Dominio | Eukaryota |
| Regno | Animalia |
| Phylum | Chordata |
| Classe | Sauropsida |
| Superordine | Dinosauria |
| Ordine | † Ornithischia |
| Sottordine | † Stegosauria |
| Famiglia | Stegosauridae Marsh, 1880 |
| Sottogruppi | |
| |

Il clado degli Stegosauridae è composto dai generi Stegosaurus, Dacentrurus, Miragaia, Loricatosaurus e Kentrosaurus, quest'ultimo considerato alla base del clado.[10] Gli stegosauridi come tutti gli animali appartenenti a Stegosauria erano grandi erbivori quadrupedi, caratterizzati da grandi placche ossee sul dorso e numerose spine ossee sulla parte finale della coda, chiamato thagomizer, che usavano come arma offensiva. Queste grandi placche sottili si ergevano erette allineate parasagittalmente dal collo verso la fine della coda, dove venivano rimpiazzate dagli spunzoni ossei.[7][9] Sebbene nel tempo siano state elaborate diverse teorie sulla funzione delle placche ossee, come la termoregolazione e il loro uso come display sessuale, una recente ricerca istologica ontogenetica sulle piastre e sulle punte indicherebbe che le piastre avessero diverse funzioni in base all'età dell'animale. Le punte terminali sulla coda, sarebbero state usate soprattutto dai vecchi adulti come arma di difesa.[11] Tuttavia, la teoria sulla funzione delle piastre e degli spunzoni nelle diverse fasi di vita, rimane ancora una questione in dibattito.
Gli stegosauridi si distinguono dagli huayangosauridi in quanto i primi hanno perso i denti premascellari e le placche laterali al tronco.[12] Inoltre, al contrario degli stegosauridi, gli huayangosauridi hanno lunghi teschi stretti e arti posteriori più lunghi rispetto agli arti anteriori.[9] Tuttavia, queste due caratteristiche non sono diagnostiche di stegosauridae perché possono essere presenti anche nei non-stegosauridi diversi da huayangosauridae.[6]
Descrizione
modificaCaratteristiche generali
modificaI resti craniali degli stegosauridi, così come tutti gli stegosauria, sono estremamente rari.[12] Quindi, una parte significativa della conoscenza della loro morfologia cranica viene da pochi teschi completi di Stegosaurus e resti frammentari di altri stegosauridi. In generale, gli stegosauridi hanno musi proporzionalmente lunghi, bassi e stretti con una mandibola profonda, rispetto a quella di Huayangosaurus. Gli stegosauridi mancano anche dei denti intermascellari.[12]
Tra gli stegosauridi, solo Kentrosaurus aveva un paio di spine ossee parascapulari, che sporgevano posteriormente nella parte inferiore delle piastre delle spalle. Queste spine sono lunghe, arrotondate in vista laterale, dalla forma a virgola e hanno una base allargata.[4] Si pensava che anche il Loricatosaurus avesse spine parascapulari, ma Maidment et al. (2008)[4] ha osservato che i campioni scoperti, tra cui è descritta una colonna vertebrale, ha una morfologia completamente diversa da quella dei campioni che possiedono spine parascapulari, suggerendo invece che le spine ritrovate in Loricatosaurus fossero in realtà spine della coda frammentarie. La vera e propria assenza di spine parascapulari negli altri stegosauridi è considerata una perdita secondaria dal momento che molti stegosauri basali come Gigantspinosaurus e Huayangosaurus le possedevano.[4]
Durante la loro evoluzione gli stegosauridi hanno perso le righe di osteodermi laterali che correvano longitudinalmente su entrambi i lati del tronco di Huayangosaurids e degli Ankylosauri, indicando un'altra perdita secondaria di un carattere plesiomorfico.[12] Tuttavia, l'assenza di scudi laterali e denti premascellari non sono specificamente diagnostici degli stegosauridi, poiché queste caratteristiche sono presenti anche in altri stegosauri non-huayangosauridi, cui le relazioni filogenetiche all'interno Stegosauria non sono chiare.[4][12]
Placche e spine
modifica
Tutti i generi di stegosauridi presentano le caratteristiche placche ossee dorsali, che non sono altro che osteodermi dorsali modificati, e spine ossee. Le piastre degli stegosauridi hanno una base spessa e centrale, ma sono trasversalmente sottili. In particolare in Stegosaurus le piastre diventano molto consistenti e sottili. Si trovano in varie dimensioni lungo il dorso, divenendo più grandi, alte e spesse soprattutto nel centro della schiena. La disposizione di queste piastre dorsali parasagittali è stata ampiamente discussa in passato. Marsh suggerì una singola fila mediana di piastre che partiva dal cranio percorrendo l'asse longitudinale[13] mentre Lull sostenne una disposizione accoppiata bilaterale.[14] L'attuale consenso scientifico risiede nella disposizione proposta da Gilmore, con due file parasagittali alternate sfalsate, teoria proposta dopo la scoperta di uno scheletro quasi completo conservato con questo arrangiamento nella roccia.[15] Inoltre, non esistono due piastre che condividano la stessa dimensione e forma, rendendo l'idea di due righe bilateralmente accoppiate ancora meno probabile. Le piastre hanno di solito scanalature vascolari distinte sulle loro superfici laterali, suggerendo la presenza di una rete circolatoria. Gli stegosauridi hanno anche osteodermi sulla gola in forma di piccoli ossicini depressi e due paia di spine ossee allungate sulla coda.[6]

In un'analisi ontogenetica istologica sulle piastre e sulle spine di Stegosaurus, Hayashi et al. (2012)[11] hanno esaminato la struttura e la funzione delle placche e delle spine in esemplari giovani ed adulti. Essi hanno scoperto che durante l'ontogenesi, gli osteodermi dorsali sono composti da dense fibre collagene solidificate sia nel tessuto osseo solido sia nel tessuto osseo spugnoso, suggerendo che le piastre e le punte fossero formate dalla diretta mineralizzazione delle reti fibrose già esistenti nella pelle. Tuttavia, le molte caratteristiche strutturali, viste nelle spine e nelle placche degli esemplari più adulti, sono acquisite a diversi stadi dello sviluppo. Le reti vascolari estese si formano sulle piastre durante il passaggio da giovani a sub-adulti e persistono negli individui più vecchi, mentre le spine acquisiscono una corteccia di spessore con un grande canale vascolare assiale, solo negli esemplari adulti più vecchi. Hayashi et al. sostengono che la formazione di reti vascolari nei giovani adulti sostengano la crescita delle piastre. Questo avrebbe migliorato le dimensioni dell'animale, contribuendo ad attrarre potenziali compagni e scoraggiare i rivali.[11] Inoltre, la presenza di reti vascolari nelle piastre di giovani adulti indica un uso secondario delle piastre come dispositivo di termoregolazione per la dispersione di calore, in modo simile alle orecchie degli elefanti, al becco dei tucani e agli osteodermi dei coccodrilli. L'ispessimento della sezione corticale dell'osso e la compattazione delle ossa nelle spine della coda negli esemplari adulti più vecchi, suggerisce che essi venivano usati attivamente come arma di difesa, ma non fino ad uno stadio ontogeneticamente ritardo. Lo sviluppo del canale assiale negli esemplari adulti più vecchi, già presenti negli esemplari adulti più giovani facilitavano ulteriore l'allargamento delle spine, aumentando la quantità di nutrimento fornita. D'altra parte, le piastre non mostrano un grado simile di compattazione dell'osso o di ispessimento corticale indicando che non sarebbero state in grado di sopportare molto peso. Questo suggerisce che le placche non sarebbero state una grande arma di difesa.[11]
La natura protettiva delle piastre dorsali era già stata messa in dubbio in passato da Davitashvili (1961), che aveva notato che la stretta posizione dorsale delle piastre lasciava i fianchi dell'animale vulnerabili. Poiché la forma delle piastre e delle spine variavano da specie a specie, Davitashvili ha suggerito che le placche fossero invece importanti per il riconoscimento intraspecifico e come display per la selezione sessuale.[6] Ciò è confermato dalle osservazioni di Spassov (1982), che notò che le piastre erano disposte proprio in modo che fossero ben visibili lateralmente durante il comportamento agonistico non aggressivo, anziché in una posizione aggressiva frontalmente.[16]
La scoperta di un'impressione di pelle che copriva le piastre dorsali implica tutte le possibili funzioni delle piastre degli stegosauri. Christiansen e Tschopp (2010)[17] hanno scoperto che la pelle sulle piastre era liscia, con lunghe scanalature parallele poco profonde che indicano una struttura cheratinosa che ricopriva le piastre. L'aggiunta di beta-cheratina, una proteina della famiglia della cheratina, consentirebbe infatti alle piastre di sopportare una maggiore pressione e peso, suggerendo che potrebbero effettivamente essere utilizzate come difesa attiva. Un rivestimento corneo darebbe anche una maggiore superficie per le piastre per essere utilizza come strutture espositive d'accoppiamento, che potrebbero essere potenzialmente molto colorate, come i becchi degli uccelli moderni. Allo stesso tempo però, il rivestimento di cheratina impedisce l'uso delle placche nella termoregolazione, in quanto il rivestimento di cheratina renderebbe il trasferimento di calore dal tessuto osseo altamente inefficace.[17]
Postura
modificaUn'articolazione digitale e la manipolazione delle scansioni digitali del materiale di un esemplare di Kentrosaurus lascia dedurre che gli stegosauridi avevano una postura quadrupede eretta dritta su tutte e quattro le zampe, come quella della maggior parte dei mammiferi, durante la loro locomozione abituale, mentre utilizzavano una posa più da coccodrillo quando attuavano un comportamento difensivo allargando le zampe per mantenere la stabilità nei movimenti della coda. La posa da sdraiato permetterebbe loro di tollerare le grandi forze laterali utilizzate per far oscillare la coda contro i predatori.[18]
Classificazione
modifica
Nel 1877, Othniel Marsh scoprì e descrisse Stegosaurus armatus, da cui nacque il nome della famiglia di Stegosauridae, eretta nel 1880.[10] Rispetto agli Huayangosauridae, le sinapomorfie notevoli di stegosauridae includono un grande antitrochantere (processo supracetabulare) sull'ileo, un lungo processo prepubico e un lungo femore rispetto alla lunghezza dell'omero.[19] Inoltre, le costole sacrali degli stegosauridi formano una T in sezione parasagittale[6] e le vertebre dorsali hanno un arco neurale allungato.[10]
Gli Stegosauridi originariamente descritti con i generi Wuerhosaurus ed Hesperosaurus sono oggi invece indicati come delle specie del genere Stegosaurus, rispettivamente come le specie S. homheni e S. mjosi, da Maidment et al. (2010), sebbene non tutti gli studiosi siano d'accordo.[10] Inoltre, molti esemplari di Stegosaurus che in precedenza erano ritenute specie diverse, all'interno del genere, sono stati tutte assegnate a Stegosaurus armatus. Questa riclassificazione dei campioni si è verificato dopo aver constatato che tutte le differenze tra i campioni possono essere spiegati con le variazioni intraspecifiche.[10]
La struttura dell'albero genealogico degli stegosauridi è stata discussa per lungo tempo. Tuttavia, in due delle più vaste analisi cladistiche sugli Stegosauri condotta fino ad oggi, sia Mateus et al. (2009)[20] sia Maidment (2010)[10] presentano relazioni filogenetiche simili all'interno stegosauridae (vedi sotto). Buona parte della filogenesi è stata risolta con la scoperta dello stegosauride dal collo lungo Miragaia, come sister taxon di Dacentrurus, formando il clado dei Dacentrurinae. Mateus et al. ha notato che in entrambe le proiezioni delle costole cervicali posteriori e il processo pubico anteriore è dorsoventralmente profondo e porta una proiezione dorsale alla sua estremità anteriore in vista laterale. I punti della loro filogenesi punti in comune con il genere Stegosaurus, sostenuto dalla presenza condivisa di un postzygapophyside sulle vertebre cervicali che sono processi allungati posteriormente alla faccetta posteriore del centrum. Il Dacentrurus era stato precedentemente considerato uno stegosauride basale, ma la scoperta e l'osservazione di Miragaia ha suggerito che sia un taxon derivato.[20]
| Stegosauria |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Paleobiologia
modificaDimorfismo sessuale
modifica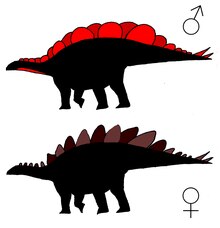
Ci sono stati diversi ritrovamenti di possibile dimorfismo sessuale negli stegosauridi. Saitta et al.(2015)[21] notò che le piastre di Stegosaurus potevano essere suddivise in due categorie: alcune placche erano basse e larghe e dalla forma vagamente ovale, mentre le seconde avevano una superficie inferiore del 45%, ed erano più alte e strette. Considerando che le piastre dorsali molto probabilmente funzionavano come strutture espositive e che la forma bassa e ovale conferiva una maggiore visualizzazione, Saitta ha assegnato questo tipo di placca ai maschi mentre quelle più strette e appuntite alle femmine.
Kevin Padian, un paleontologo dell'Università della California, Berkeley, ha osservato che non tutti gli esemplari presi in considerazione da Saitta era pienamente cresciuti, affermando quindi che tale morfologia potrebbe semplicemente essere data dall'età dell'animale.[22]
Tuttavia un'altra possibile interpretazione di dimorfismo sessuale negli stegosauridi Kentrosaurus, Dacentrurus e Stegosaurus, sarebbe data dalla presenza di tre costole sacrali in più nelle femmine.[6]
Alimentazione
modificaAl fine di comprendere le abitudini alimentari degli stegosauridi, Reichel (2010)[23] ha creato un modello 3-D dei denti di uno Stegosaurus, utilizzando il software ZBrush. Il modello ha rivelato che la forza del morso di uno Stegosaurus era significativamente più debole di quella del Labrador, di un lupo o di un uomo. La scoperta suggerisce che questi dinosauri sarebbero stati in grado di rompere i rami e le foglie più piccole grazie ai loro denti, ma non sarebbero stati in grado di mordere e strappare un oggetto di 12 mm o più di diametro. Parrish et al. (2004)[24] nella sua descrizione della flora del Giurassico della Formazione Morrison, supportava questa constatazione. La flora durante questo periodo di tempo era dominata da piccoli piante erbacee stagionali dalla crescita rapida, che uno stegosauride avrebbe consumato facilmente, se la ricostruzione di Reichel fosse accurata.[23]
Mallison (2010)[18] suggerì che il Kentrosaurus avrebbe potuto occasionalmente adottare una posizione tripode, ergendosi sulle zampe posteriori e puntellandosi al terreno con la coda per raggiungere il cibo che stava più in alto. Ciò ha sfidato l'opinione pubblica che stegosauridi potessero cibarsi di vegetali solo ad un metro dal suolo a causa delle teste piccole e dal collo e arti anteriori corti, dal momento che la posizione tripode avrebbe anche dato loro l'accesso a giovani alberi e cespugli alti.
Un altro indizio che suggeriscono che alcuni stegosauridi potevano nutrirsi di vegetazione più alta è data dalla scoperta dello stegosauride dal collo lungo Miragaia longicollum. Il collo di questo dinosauro aveva almeno 17 vertebre cervicali, dovute alla trasformazione di vertebre toraciche in vertebre cervicali. Questo procedimento era già stato sviluppato dai sauropodi per lo stesso motivo.[20]
Note
modifica- ^ T.A. Tumanova e V.R. Alifanov, First Record of Stegosaur (Ornithischia, Dinosauria) from the Aptian-Albian of Mongolia, in Paleontological Journal, vol. 52, n. 14, 2018, pp. 1771–1779.
- ^ F. Costa e O. Mateus, Dacentrurine stegosaurs (Dinosauria): A new specimen of Miragaia longicollum from the Late Jurassic of Portugal resolves taxonomical validity and shows the occurrence of the clade in North America, in PLOS ONE, vol. 14, n. 11, 2019, pp. e0224263.
- ^ O. Zafaty, M. Oukassou, F. Riguetti, J. Company, S. Bendrioua, R. Tabuce, A. Charrière e X. Pereda-Suberbiola, A new stegosaurian dinosaur (Ornithischia: Thyreophora) with a remarkable dermal armour from the Middle Jurassic of North Africa, in Gondwana Research, 2024.
- ^ a b c d e Susannah C. R. Maidment, David B. Norman, Paul M. Barrett e Paul Upchurch, Systematics and phylogeny of Stegosauria (Dinosauria: Ornithischia), in Journal of Systematic Palaeontology, vol. 6, n. 4, 1º gennaio 2008, pp. 367–407, DOI:10.1017/S1477201908002459, ISSN 1477-2019.
- ^ O.W.M. Rauhut, J.L. Carballido e D. Pol, First Osteological Record of a Stegosaur (Dinosauria, Ornithischia) from the Upper Jurassic of South America, in Journal of Vertebrate Paleontology, vol. 40, n. 6, 2021, pp. e1862133.
- ^ a b c d e f David B. Weishampel, Peter Dodson, Halszka Osmólska. The Dinosauria (2nd ed.). Berkeley: University of California Press.
- ^ a b (EN) Jean-Paul Billon-Bruyat e Daniel Marty, Preface: Symposium on Stegosauria proceedings, in Swiss Journal of Geosciences, vol. 103, n. 2, 4 settembre 2010, pp. 139–141, DOI:10.1007/s00015-010-0027-z, ISSN 1661-8726.
- ^ Ulansky R. E., 2014. Dinosaurs Classification. Basal Thyreophora & Stegosauria. Dinologia,
- ^ a b c (EN) Paul C. Sereno, The Evolution of Dinosaurs, in Science, vol. 284, n. 5423, 25 giugno 1999, pp. 2137–2147, DOI:10.1126/science.284.5423.2137, ISSN 0036-8075, PMID 10381873.
- ^ a b c d e f (EN) Susannah C. R. Maidment, Stegosauria: a historical review of the body fossil record and phylogenetic relationships, in Swiss Journal of Geosciences, vol. 103, n. 2, 7 settembre 2010, pp. 199–210, DOI:10.1007/s00015-010-0023-3, ISSN 1661-8726.
- ^ a b c d (EN) Shoji Hayashi, Kenneth Carpenter, Mahito Watabe e Lorrie A. McWHINNEY, Ontogenetic histology of Stegosaurus plates and spikes, in Palaeontology, vol. 55, n. 1, 1º gennaio 2012, pp. 145–161, DOI:10.1111/j.1475-4983.2011.01122.x, ISSN 1475-4983.
- ^ a b c d e Sereno, Paul C., and Dong Zhimin. "The Skull of the Basal Stegosaur Huayangosaurus Taibaii and a Cladistic Diagnosis of Stegosauria." Journal of Vertebrate Paleontology 12, no. 3 (1992): 318-43. http://www.jstor.org/stable/4523456.
- ^ Marsh, O. C. (1891). Restoration of Stegosaurus. American Journal of Science, 3rd series, 42, 179–182.
- ^ Lull, R. S. (1910a). Stegosaurus ungulatus Marsh, recently mounted at the Peabody Museum of Yale University. American Journal of Science, 4th series, 30, 361–377
- ^ Gilmore, C. W. (1914). Osteology of the armored Dinosauria in the United States National Museum, with special reference to the genus Stegosaurus. United States National Museum Bulletin, 89, 1–143.
- ^ Spassov, N. B. (1982). The ‘‘bizarre’’ dorsal plates of stegosaurs: ethological approach. Comptes rendus de l’academie bulgare des Sciences, 35, 367–370.
- ^ a b (EN) Nicolai A. Christiansen e Emanuel Tschopp, Exceptional stegosaur integument impressions from the Upper Jurassic Morrison Formation of Wyoming, in Swiss Journal of Geosciences, vol. 103, n. 2, 7 settembre 2010, pp. 163–171, DOI:10.1007/s00015-010-0026-0, ISSN 1661-8726.
- ^ a b (EN) Heinrich Mallison, CAD assessment of the posture and range of motion of Kentrosaurus aethiopicus Hennig 1915, in Swiss Journal of Geosciences, vol. 103, n. 2, 7 settembre 2010, pp. 211–233, DOI:10.1007/s00015-010-0024-2, ISSN 1661-8726.
- ^ David E. Fastovsky , David B. Weishampel. The Evolution and Extinction of the Dinosaurs. p. 123
- ^ a b c (EN) Octávio Mateus, Susannah C. R. Maidment e Nicolai A. Christiansen, A new long-necked ‘sauropod-mimic’ stegosaur and the evolution of the plated dinosaurs, in Proceedings of the Royal Society of London B: Biological Sciences, vol. 276, n. 1663, 22 maggio 2009, pp. 1815–1821, DOI:10.1098/rspb.2008.1909, ISSN 0962-8452, PMC 2674496, PMID 19324778.
- ^ Evan Thomas Saitta, Evidence for Sexual Dimorphism in the Plated Dinosaur Stegosaurus mjosi (Ornithischia, Stegosauria) from the Morrison Formation (Upper Jurassic) of Western USA, in PLOS ONE, vol. 10, n. 4, 22 aprile 2015, pp. e0123503, DOI:10.1371/journal.pone.0123503, ISSN 1932-6203, PMC 4406738, PMID 25901727.
- ^ Dino 'sexing' study slammed by critics, su Science | AAAS, 22 aprile 2015. URL consultato il 4 giugno 2016.
- ^ a b (EN) Miriam Reichel, A model for the bite mechanics in the herbivorous dinosaur Stegosaurus (Ornithischia, Stegosauridae), in Swiss Journal of Geosciences, vol. 103, n. 2, 31 agosto 2010, pp. 235–240, DOI:10.1007/s00015-010-0025-1, ISSN 1661-8726.
- ^ Judith Totman Parrish, Fred Peterson e Christine E Turner, Jurassic "savannah"—plant taphonomy and climate of the Morrison Formation (Upper Jurassic, Western USA), in Sedimentary Geology, Reconstruction of the Extinct Ecosystem of the Upper Jurassic Morrison Formation, vol. 167, 3–4, 15 maggio 2004, pp. 137–162, DOI:10.1016/j.sedgeo.2004.01.004.
Altri progetti
modifica Wikimedia Commons contiene immagini o altri file su Stegosauridae
Wikimedia Commons contiene immagini o altri file su Stegosauridae Wikispecies contiene informazioni su Stegosauridae
Wikispecies contiene informazioni su Stegosauridae
Collegamenti esterni
modifica- (EN) Stegosauridae, su Fossilworks.org.