Le informazioni riportate non sono consigli medici e potrebbero non essere accurate. I contenuti hanno solo fine illustrativo e non sostituiscono il parere medico: leggi le avvertenze.
Un virus (dal latinovīrus, "veleno") è un'entità biologica con caratteristiche di simbionte o parassita obbligato, in quanto si replica esclusivamente all'interno delle cellule degli organismi. I virus possono infettare tutte le forme di vita, dagli animali, alle piante, ai microrganismi (compresi altri agenti infettanti come i batteri) e anche altri virus.[1][2] Molte specie di virus convivono all'interno di sistemi viventi ospiti in modo simbiotico e gli individui di ogni specie animale, compreso l'essere umano, ospitano normalmente un elevato numero di specie virali simbionti, che formano una popolazione detta viroma.
I virus nella loro forma isolata, detta virione, sono entità biologiche ultramicroscopiche, cioè invisibili al microscopio ottico, con un ordine di grandezza variabile dalla decina al centinaio di nanometri, mentre un microscopio ottico raggiunge al massimo i 0,2 μm. (un micrometro è un milionesimo di metro, mentre un nanometro è un miliardesimo di metro, cioè mille volte più piccolo). Sono quindi di dimensioni più piccole dei microrganismi e sono composti da filamenti di DNA o RNA racchiusi all'interno di un involucro proteico, detto capside, che può avere forme diverse; possono inoltre contenere enzimi ed essere circondati da un ulteriore involucro esterno, formato da uno strato lipidico e glicoproteine simile alle membrane cellulari, detto pericapside. Tra i due strati protettivi possono esservi proteine specifiche.[3] Le forme dei virus possono variare notevolmente, ma alcune sono più comuni: tra queste vi sono le tipologie sferiche, poliedriche, elicoidali, filiformi, amorfe o composite.
La scoperta dei virus risale al 1892, quando Dmitrij Ivanovskij descrisse in un articolo un agente patogeno non batterico in grado di infettare le piante di tabacco, da cui nel 1898[4]Martinus Beijerinck scoprì il virus del mosaico del tabacco. Da allora, circa 5 000 specie di virus sono state descritte in dettaglio,[5] anche se si ritiene che ve ne siano milioni.[6] I virus si trovano in quasi tutti gli ecosistemi della Terra e rappresentano l'entità biologica più abbondante in assoluto.[7][8] La disciplina che si occupa dello studio dei virus è nota come virologia, una sub-specialità della microbiologia.
I virus hanno diverse caratteristiche degli esseri viventi poiché possiedono un genoma, si riproducono e si evolvono attraverso la selezione naturale. Tuttavia, secondo la maggior parte degli autori, pur contenendo DNA o RNA, non sono considerabili esseri viventi in quanto non organizzati in cellule, non dotati di vita autonoma e non in grado di replicarsi autonomamente.[9][10] Poiché ne possiedono solo alcune caratteristiche sono stati descritti anche come "organismi ai margini della vita"[11] o come "replicatori".[12]
Quando non si trovano nella fase dell'infezione o all'interno di una cellula infetta, i virus esistono in forma di particelle indipendenti e inattive. Queste particelle virali, note anche come virioni, sono costituite da due o tre parti: (I) il materiale genetico costituito da DNA o RNA, lunghe molecole che trasportano le informazioni genetiche; (II) un rivestimento proteico, chiamato capside, che circonda e protegge il materiale genetico; e in alcuni casi (III) un involucro esterno formato da uno strato di lipidi, che circonda il rivestimento proteico, detto pericapside. Non tutti i virus sono dotati di questo secondo involucro (i virus delle piante ad esempio, in genere ne sono privi). I virioni possono avere forme semplici, elicoidali e icosaedriche, ma anche architetture più complesse. La maggior parte dei virus possiede virioni che sono troppo piccoli per essere visti con un microscopio ottico. In media il virione ha una dimensione di circa un centesimo della dimensione media di un batterio.
I virus hanno bisogno di un ospite (una cellula, eventualmente parte di un organismo pluricellulare, o un altro sistema biologico) per replicarsi, ma hanno anche la necessità di trasmettersi da un ospite all'altro. Il processo può avvenire in molti modi: quelli delle piante hanno generalmente necessità di un vettore, cioè un organismo intermedio mobile, pertanto spesso vengono trasmessi da pianta a pianta da insetti che si nutrono della loro linfa (ad esempio gli afidi). Lo stesso meccanismo può avvenire negli animali a opera di parassiti succhiatori di sangue. La maggioranza dei virus non vegetali ha però capacità di trasmettersi senza il contributo di altre specie vettori. I virus influenzali ad esempio si diffondono direttamente da un ospite all'altro attraverso le esalazioni respiratorie, e ricevono particolare diffusione tramite fenomeni come tosse e starnuti. I norovirus e i rotavirus, comuni cause di gastroenterite virale, sono trasmessi per via oro-fecale, vengono trasportati e depositati in luoghi esterni (come oggetti, alimenti o fonti d'acqua) tramite contatto, e possono entrare nel corpo con cibo e bevande contaminate. L'HIV è uno dei numerosi virus trasmessi attraverso il contatto sessuale o dallo scambio di fluidi corporei, come il sangue. L'insieme delle tipologie di cellule che un virus può utilizzare per replicarsi, viene chiamata il suo "ospite". Si tratta sempre di cellule dotate di sistemi adatti a replicare le sequenze genetiche contenute nel virus. Inoltre, i virus devono anche essere dotati di meccanismi specializzati per riuscire a oltrepassare le membrane delle cellule ospiti. Alcuni virus sono in grado di infettare solo pochi ospiti, altri invece sono in grado di infettarne molti.[14]
Negli animali, le infezioni da virus dannosi provocano una risposta immunitaria che solitamente elimina o comunque neutralizza il virus infettante. Gli animali sono generalmente dotati di difese molto potenti nei confronti dei virus dannosi, che consistono non soltanto in meccanismi di risposta, automatica o mirata, ma anche in barriere fisiche e chimiche molto difficili da oltrepassare per i virus esterni. Le risposte immunitarie possono essere prodotte anche da vaccini, che conferiscono un'immunità artificialmente acquisita per l'infezione virale specifica. Tuttavia, alcuni virus, come quelli che causano l'AIDS e l'epatite virale, sono in grado di eludere queste risposte immunitarie e provocano infezioni croniche. Poiché i virus non hanno un metabolismo, i farmaci antibiotici non possono eliminarli e non hanno alcun effetto diretto su di essi. Sono stati osservati effetti indiretti di taluni farmaci antibiotici e antiparassitari che inibiscono meccanismi vitali delle cellule ospiti[15][16][17], ma la loro efficacia resta dubbia o marginale.
La parola "virus" deriva dalla parola latinavīrus, che significa "tossina" o "veleno".[18]
Quelle del mosaico del tabacco sono state le prime formazioni virali a essere osservate dall'uomo nel 1892, per opera dello scienziato russo Dmitrij Iosifovič Ivanovskij. Furono inizialmente chiamati "virus filtrabili" in quanto passavano attraverso filtri che trattenevano i batteri, filtri in ceramica a porosità micrometrica ma, a differenza delle semplici tossine, potevano essere trattenuti da ultrafiltri. Saranno classificati definitivamente come virus nel 1898 dal botanico olandese Martinus Willem Beijerinck il quale, usando esperimenti di filtrazione su foglie di tabacco infette, riuscì a dimostrare che il mosaico del tabacco è causato da un agente infettivo di dimensioni minori di quelle di un batterio.
La scelta del vocabolo latino vīrus causa problemi nel voler ricavarne il plurale, come si suol fare nella tassonomia (vedi animalia per animali, plantae, per piante, eccetera). Essendo una parola neutra della II declinazione e terminando in -us nei tre casi diretti (nominativo, accusativo e vocativo) è considerata irregolare. Si declina nei casi obliqui (genitivo, dativo e ablativo) con il sinonimo venēnum. Dovrebbe essere singulare tantum, ha cioè soltanto il singolare, come "riso" o "aria". In circostanze particolari potrebbe essere reso plurale, ma non ci è giunta tale forma. Lwoff, Horne e Tournier, nella loro classificazione del 1962, propongono e usano la forma vira. In ambito anglofono viene usata la forma errata virii (da vīriī), più che altro per riferirsi ai virus informatici, mentre in ambito biologico è usata la forma pluralizzata inglese viruses. Vīriī è errato poiché presuppone la parola vīrius (come radius, radiī), che non esiste. Altro plurale ipotizzabile è vīrī, che però richiederebbe come singolare la parola vir, che significa "uomo" e si rende al plurale come vĭrī.
Il termine "virione" (plurale "virioni"), coniato in francese nel 1959 nella forma "virion"[19] ed entrato nella lingua italiana nel 1983, è inoltre usato per riferirsi a una singola particella virale, stabile e infettiva, che viene rilasciata dalla cellula ed è pienamente in grado di infettare altre cellule dello stesso tipo.[20]
Nel 1892 il biologorussoDmitri Ivanovsky utilizzò questo filtro per studiare quello che oggi è conosciuto come il virus del mosaico del tabacco. I suoi esperimenti hanno dimostrato che estratti di foglie da piante di tabacco schiacciate e infettate rimangono infettivi dopo la filtrazione. Ivanovsky suggerì che l'infezione poteva essere causata da una tossina prodotta da batteri, ma non approfondì la questione.[23] Al momento si pensava che tutti gli agenti infettivi potessero essere trattenuti dai filtri e coltivati su un terreno nutritivo.[24] Nel 1898, il microbiologo olandese Martinus Beijerinck ripeté gli esperimenti e si convinse che la soluzione filtrata conteneva una nuova forma di agente infettivo.[25] Egli osservò che questo agente si moltiplicava solo nelle cellule che si stavano dividendo, ma poiché i suoi esperimenti non mostrarono che fosse fatto di particelle, lo chiamò contagium vivum fluidum (solubile germe di vita), e reintrodusse la parola virus.[23] Beijerinck sostenne che i virus fossero liquidi in natura, una teoria più tardi screditata da Wendell Meredith Stanley, che dimostrò che erano delle particelle.[23] Nello stesso anno Friedrich Loeffler e Paolo Frosch isolarono il primo virus animale, l'agente dell'afta epizootica (aphthovirus), attraverso un filtro simile.[26]
Nel XX secolo, il batteriologo inglese Frederick Twort scoprì un gruppo di virus capaci di infettare i batteri, oggi chiamati batteriofagi[24] (o comunemente fagi) e il microbiologo franco-canadese Félix d'Herelle descrisse un virus che, quando aggiunto a batteri su agar, produrrebbe aree di batteri morti. Diluì accuratamente una sospensione di questi virus e scoprì che le più alte diluizioni (concentrazioni più basse di virus), piuttosto che uccidere tutti i batteri, formavano aree discrete di organismi morti. Contando questi settori e moltiplicando per il fattore di diluizione egli riuscì a calcolare il numero di virus nella sospensione originale.[27] I fagi furono considerati come un potenziale trattamento per alcune malattie come il tifo e il colera, ma tale idea fu messa da parte grazie all'introduzione della penicillina. Lo studio di fagi, tuttavia, ha permesso di capire alcuni aspetti dell'espressione genica e un meccanismo utile per introdurre geni estranei nei batteri.
Alla fine del XIX secolo, i virus vennero classificati secondo la loro infettività, la loro capacità di essere filtrati e i requisiti per i loro ospiti viventi. Inizialmente i virus vennero coltivati solo nelle piante e negli animali. Nel 1906, Ross Granville Harrison introdusse un metodo per la coltivazione nella linfa, e, nel 1913, E. Steinhardt, C. Israeli, and R. A. Lambert utilizzarono questo metodo per allevare i virus destinati a diventare vaccini in frammenti di tessuto corneale di una cavia.[28] Nel 1928, HB Maitland e MC Maitland fecero moltiplicare i virus-vaccino in sospensioni di reni di galline. Il loro metodo incominciò a essere ampiamente adottato a partire dal 1950, quando il poliovirus venne coltivato su larga scala per la produzione del vaccino antipoliomielite.[29]
Modelli tridimensionale in scala di virus ebola (grigio) e virus batteriofago (marone)
Le prime immagini di virus sono state ottenute in seguito all'invenzione del microscopio elettronico nel 1931, grazie al lavoro degli ingegneri tedeschi Ernst Ruska e Max Knoll.[32] Nel 1935, il biochimico e virologo americano Wendell Meredith Stanley analizzò il virus del mosaico del tabacco e dimostrò che era per la maggior parte costituito da proteine.[33] Poco tempo dopo, fu possibile separare in questo virus le proteine dall'RNA.[34] Il virus del mosaico del tabacco è stato il primo a essere cristallizzato e la sua struttura è stata conseguentemente analizzata nel dettaglio. Le prime immagini a raggi X di diffrazione del virus cristallizzato, furono ottenute da Bernal e Fankuchen nel 1941. Sulla base delle loro foto, Rosalind Franklin scoprì, nel 1955, la struttura completa del virus.[35] Nello stesso anno, Heinz Fraenkel-Conrat e Robley Williams dimostrarono che il virus del mosaico del tabacco purificato e il suo rivestimento proteico sono in grado di assemblare da soli dei virus funzionali, suggerendo che questo semplice meccanismo è probabilmente stato il mezzo attraverso cui i virus vengono creati all'interno delle loro cellule ospiti.[36]
I virus sono tutti parassiti intracellulari obbligati. All'esterno delle cellule ospiti sono costituiti da un virione, formato da una capsula proteica (detta capside) contenente l'acido nucleico (DNA o RNA). I virus degli Eucarioti possono possedere anche una membrana che avvolge il capside detta peplos o pericapside. Talvolta tra il capside e il pericapside presentano un ulteriore strato proteico che prende il nome di tegumento. I virioni non possiedono metabolismo: vengono quindi trasportati passivamente finché non trovano una cellula da infettare. L'infezione di una cellula ospite richiede il legame con proteine specifiche di membrana.
Nelle cellule infettate i virus perdono la loro individualità strutturale: consistono negli acidi nucleici e nei loro prodotti che assumono il controllo di parte dell'attività biosintetica cellulare al fine di produrre nuovi virioni.
In alternativa, alcuni virus possono inserire fisicamente il loro genoma in quello dell'ospite in modo che sia replicato insieme con esso. Il genoma virale inserito in quello dell'ospite, detto provirus, riprende la sua individualità e produce nuovi virioni in caso di danneggiamento della cellula ospite.
I virus forse esistono fin dall'evoluzione delle prime cellule viventi.[41] L'origine dei virus non è tuttavia chiara poiché non formano fossili nel senso classico del termine; tecniche molecolari sono state utilizzate per confrontare il DNA o l'RNA dei virus e si sono dimostrate molto utili per indagare su come si potevano presentare anticamente.[42] Inoltre, il materiale genetico virale può occasionalmente passare nella linea germinale degli organismi ospitanti, per cui i discendenti ospitanti avranno il virus integrato nel proprio genoma. Ciò fornisce una preziosa fonte di informazioni per i virologi per poter risalire agli antichi virus che sono esistiti liberi fino a milioni di anni fa.
Vi sono tre ipotesi principali, qui di seguito descritte, che mirano a spiegare le origini dei virus.[43][44]
Questa teoria afferma che i virus potrebbero essere stati una volta delle piccole cellule parassitarie di cellule più grandi. Nel corso del tempo, i geni non richiesti dalla loro natura parassitaria sono andati persi. I batteri dei generi Rickettsia e Clamidia sono le cellule che, come i virus, sono in grado di riprodursi solo all'interno di cellule ospiti viventi. Essi prestano sostegno a questa ipotesi per il fatto che la loro dipendenza da parassitismo probabilmente causò loro la perdita dei geni che gli potevano permettere di sopravvivere al di fuori di una cellula. Questa ipotesi è chiamata anche di degenerazione[45][46] o di riduzione.[47]
Questa, talvolta chiamata ipotesi del vagabondaggio[45][48] o ipotesi di fuga,[47] consiste nel credere che alcuni virus si siano evoluti da frammenti di DNA o RNA che sono "sfuggiti" da geni di un organismo più grande. Il DNA fuggito sarebbe potuto provenire da plasmidi (frammenti di DNA che possono muoversi tra le cellule) o trasposoni/retrotrasposoni (molecole di DNA che si replicano e si muovono da diverse posizioni all'interno dei geni della cellula).[49] Una volta chiamati "geni che saltano", i trasposoni sono esempi di elementi genetici mobili che potrebbero essere l'origine di alcuni virus perché hanno delle sequenze analoghe. Essi sono stati scoperti nel mais da Barbara McClintock nel 1950.[50]
Questa è chiamata anche ipotesi del primo virus[47] e propone che i virus si siano evoluti da molecole complesse di proteine e acidi nucleici contemporaneamente alla prima apparizione sulla Terra della cellula e sarebbero rimasti dipendenti dalla vita cellulare per miliardi di anni. I viroidi sono molecole di RNA che non sono classificati come virus poiché mancano di un rivestimento proteico. Tuttavia, essi hanno caratteristiche che sono comuni a diversi virus e sono spesso chiamati agenti subvirali.[51] I viroidi sono importanti patogeni delle piante.[52] Questi non codificano proteine ma interagiscono con la cellula ospite e utilizzano le sue strutture per replicarsi.[53] Il virus dell'epatite delta degli esseri umani ha un genoma RNA simile ai viroidi, ma è dotato di un rivestimento proteico derivato dal virus dell'epatite B e non può produrne uno suo. È, quindi, un virus difettoso. Sebbene il genoma del virus dell'epatite delta possa replicarsi indipendentemente all'interno di una cellula ospite, esso richiede la presenza del virus dell'epatite B che gli fornisca un rivestimento proteico in modo che possa essere trasmesso alle nuove cellule.[54] In modo analogo, il virofago Sputnik dipende dai mimivirus, e infetta il protozooAcanthamoeba castellanii.[55] Questi virus, che dipendono dalla presenza di altre specie di virus nella cellula ospite, sono chiamati "satelliti" e possono rappresentare intermedi evolutivi dei viroidi e dei virus.[56][57]
In passato, vi sono stati problemi con tutte queste ipotesi: l'ipotesi regressiva non spiega il motivo per cui anche il più piccolo dei parassiti cellulari non assomigli ai virus in alcun modo. L'ipotesi della fuga non spiega la presenza di capsidi complessi e altre strutture di particelle virali. L'ipotesi del primo virus viola la stessa definizione di virus che richiede una cellula ospite.[47]
I virus sono ora riconosciuti come molto antichi e con una data d'origine antecedente alla divergenza della vita nei tre domini.[58] Questa scoperta ha portato i virologi moderni a riconsiderare e rivalutare le tre ipotesi classiche.[58].
L'evidenza di un mondo ancestrale delle cellule RNA[59] e le analisi al computer di sequenze di DNA virale stanno fornendo una migliore comprensione delle relazioni evolutive tra i diversi virus e potranno aiutare a identificare gli antenati dei virus moderni. Queste analisi non hanno potuto dimostrare quale delle ipotesi formulate sia più corretta[59], tuttavia sembra improbabile che tutti i virus conosciuti abbiano un antenato comune e probabilmente molti virus sono nati in passato con diversi meccanismi.[60]
Vi è una divergenza di opinioni sulla questione se i virus siano una forma di vita o strutture organiche che interagiscono con gli organismi viventi. Sono stati descritti da un autore come "organismi ai margini della vita",[11] dal momento che assomigliano agli organismi che possiedono i geni e si evolvono per selezione naturale[61] e si riproducono tramite la creazione di più copie di loro stessi attraverso un meccanismo di auto-assemblaggio il che li farebbe anche definire come "creatori di diversità"[62]. Nonostante possiedano geni, essi non hanno una struttura cellulare, aspetto che viene spesso visto come l'unità base della vita. I virus non hanno nemmeno un proprio metabolismo e necessitano della cellula ospite per realizzare nuovi prodotti. Essi, pertanto, non possono naturalmente riprodursi al di fuori di una cellula ospite.[63] Tuttavia specie batteriche come le rickettsie e le clamidie sono considerati organismi viventi nonostante abbiano la stessa limitazione.[64][65] Forme di vita accettate come tali utilizzano la divisione cellulare per riprodursi, mentre i virus si assemblano spontaneamente all'interno delle cellule. Si differenziano dalla crescita autonoma dei cristalli per il fatto che ereditano mutazioni genetiche e sono soggetti alla selezione naturale. L'auto-assemblaggio dei virus nelle cellule ospiti comporta implicazioni per lo studio dell'origine della vita, in quanto conferisce ulteriore credito all'ipotesi che la vita possa aver avuto origine da molecole organiche auto-assemblate.[66]
Diagramma di come un capside è in grado di costruire molteplici copie da solo due proteine
I virus mostrano una grande diversità di forme e dimensioni, chiamate morfologie. In generale, i virus sono molto più piccoli dei batteri. La maggior parte dei virus che sono stati studiati hanno un diametro tra 20 e 300 nanometri. Alcuni filovirus arrivano fino a una lunghezza totale a 1400 nm, ma il loro diametro è solo di circa 80 nm.[67] La maggior parte dei virus non possono essere visti con un microscopio ottico e dunque per visualizzare i virioni è necessario utilizzare il microscopio elettronico.[68] Per aumentare il contrasto tra i virus e lo sfondo, vengono utilizzati dei "coloranti" densi di elettroni. Si tratta di soluzioni di sali di metalli pesanti, come il tungsteno, che disperdono gli elettroni. Quando i virioni vengono rivestiti con questa macchia (colorazione positiva), i piccoli dettagli vengono oscurati. La colorazione negativa supera questo problema colorando solo lo sfondo.[69]
Struttura icosaedrica
Una particella virale completa, nota come un virione, è costituita da acido nucleico circondato da un rivestimento protettivo formato da proteine chiamato capside. Questo è realizzato da subunità proteiche identiche chiamati capsomeri.[70] I virus possono avere una "sacca" lipidica più esterna derivata dalla membrana della cellula ospite, detta pericapside.
Il capside è costituito da proteine codificate dal genoma virale e la sua forma serve come base per la distinzione morfologica.[71][72] Perché le subunità proteiche virali codificate si possano auto-assemblare per formare il capside, in generale si richiede la presenza del genoma virale. Virus complessi codificano proteine che aiutano la costruzione del loro capside. Le proteine associate all'acido nucleico sono note come nucleoproteine e l'associazione di proteine del capside virale con l'acido nucleico virale è chiamato nucleocapside. Il capside e l'intera struttura del virus possono essere fisicamente sondati attraverso il microscopio a forza atomica.[73][74] In generale, i virus si possono distinguere in cinque tipologie morfologiche:
Questi virus sono composti da un unico tipo di capsomero accatastato intorno a un asse centrale, formando una struttura elicoidale, che può avere una cavità centrale. Questa disposizione comporta virioni a forma di bastoncello o filamentosi: questi possono essere brevi ed estremamente rigidi o lunghi e molto flessibili. Il materiale genetico, in generale, RNA a singolo filamento, ma in alcuni casi anche DNA a singolo filamento, è vincolato nell'elica proteica dalle interazioni tra l'acido nucleico caricato negativamente e le cariche positive poste sulla proteina. In generale, la lunghezza di un capside elicoidale è correlato alla lunghezza dell'acido nucleico in esso contenuto e il diametro dipende dalla dimensione e disposizione dei capsomeri. Il virus del mosaico del tabacco e il virus della rabbia sono esempi ben analizzati di virus elicoidale.[75]
Esempi di virus a geometria icosaedrica: A. Virus senza membrana; B. Virus con membrana.
La maggior parte dei virus che colpiscono gli animali sono di morfologia icosaedrica o quasi sferica con simmetria icosaedrica. Un icosaedro regolare è un modo ottimale per formare un guscio chiuso da sub-unità identiche. Il numero minimo di capsomeri identici richiesto è dodici, ciascuno composto da cinque subunità identiche. Molti virus, come il rotavirus, possiedono più di dodici capsomeri e sembrano sferici pur mantenendo questa simmetria. I capsomeri agli apici sono circondati da altri cinque capsomeri e sono chiamati pentoni. I capsomeri sulle facce triangolari sono circondati da altri sei e sono chiamati esoni.[76] Gli esoni sono essenzialmente pianeggianti, mentre i pentoni che formano i 12 vertici sono curvilinei. La stessa proteina può agire come subunità sia nei pentoni sia negli esoni o possono essere composti da proteine differenti.
Alcuni virus hanno una forma a capsula allungata, costituita da una struttura cilindrica chiusa alle estremità da due calotte, che conferisce al virione una forma ovoidale che ricorda un dirigibile o un pallone da rugby. Si tratta in effetti di una struttura a icosaedro allungata di circa cinque volte lungo l'asse, ed è tipica delle teste dei batteriofagi.[77]
Modello scientificamente accurato a livello atomico della struttura esterna del virus SARS-Cov2. Il virione è incapsulato e costellato di proteine. Alcuni generi di virus sono avvolti da un involucro - detto pericapside - che ha costituzione lipidica ed è simile per struttura alle membrane cellulari. Esso viene costruito a partire da una membrana della cellula ospite infettata, la membrana esterna che circonda la cellula o le membrane interne come la membrana nucleare o il reticolo endoplasmatico, a seconda del meccanismo specifico del virus, e ne costituisce una forma modificata. Il virione ottiene così un doppio strato lipidico esterno analogo a una membrana cellulare, noto come pericapside o involucro virale, che dà alla particella una forma tondeggiante o irregolare. Questa membrana è costellata da proteine di vario tipo, codificate dal genoma virale e dal genoma ospite; la membrana lipidica stessa e qualsiasi carboidrato presente provengono interamente dall'ospite. L'involucro e le sue proteine possono agire come "chiavi" per penetrare all'interno delle cellule ospiti. Questa tipologia è molto diffusa tra i virus animali. Il virus influenzale e quello dell'HIV usano questa strategia, così come pure il virus sinciziale e quello della varicella, entrambi caratterizzati da un involucro irregolare. L'infettività della maggior parte di questi virus dipende dal loro involucro.[78]
Questi virus possiedono un capside che non è né puramente elicoidale né puramente icosaedrico e che può possedere strutture extra come code proteiche o una parete esterna complessa. Alcuni batteriofagi, come, il batteriofago t4 hanno una struttura complessa costituita da una testa icosaedrica associata a una coda elicoidale, che può avere una piastra di base esagonale con una coda proteica sporgente. Tale struttura a coda agisce come una siringa molecolare, attaccandosi al batterio ospite e per poi iniettare il genoma virale nella cellula.[79]
Pandoravirus salinus osservato al microscopio elettronicoI poxvirus sono virus grandi e complessi dotati di una morfologia inusuale. Il genoma virale è associato con le proteine all'interno di una struttura centrale del disco, conosciuta come nucleoide. Il nucleoide è circondato da una membrana e due corpi laterali dalla funzione sconosciuta. Il virus ha un involucro esterno con uno strato molto spesso di proteine costellato sulla sua superficie. L'intero virione è leggermente pleimorfico, dalla forma ovoidale.[80] Il mimivirus è uno dei più grandi virus caratterizzati, con un diametro del capside di 400 nm. I filamenti di proteine, si proiettano di 100 nm dalla superficie. Al microscopio elettronico, il capside appare esagonale, quindi è probabilmente icosaedrico.[81] Nel 2011, i ricercatori hanno scoperto il più grande virus fino allora conosciuto in campioni di acqua prelevati dal fondo dell'oceano Pacifico al largo della costa di Las Cruces, in Cile. Provvisoriamente chiamato Megavirus chilensis, può essere visto con un microscopio ottico di base.[82] Nel 2013, il genere pandoravirus è stato scoperto in Cile e in Australia e possiede un genoma circa due volte più grande del megavirus chilensis e del mimivirus.[83] Questi generi di virus differiscono enormemente tra loro per forma e genoma, ma condividono la caratteristica di infettare le amebe, e si sospetta possano essere molto comuni.
Alcuni virus in grado di infettare gli Archaea hanno strutture complesse che sono estranee a qualsiasi altra forma di virus, con una grande varietà di forme insolite, che vanno da strutture a forma di fuso, ai virus che assomigliano ad aste uncinate o anche a bottiglie. Altri virus archeali assomigliano alla coda dei batteriofagi e possono avere strutture multiple di coda.[84]
Un'enorme varietà di strutture genomiche possono essere riscontrate tra le specie virali; come gruppo, essi contengono una diversità genomica strutturale maggiore rispetto alle piante, agli animali, agli archeobatteri o ai batteri. Vi sono milioni di diverse specie di virus,[6] anche se solo circa 5 000 specie sono stati descritte in dettaglio.[5] A settembre 2015, il database NCBI dei genoma dei virus contava più di 75 000 sequenze genomiche complete[85], ma senza dubbio ve ne sono molte di più ancora da scoprire.[86]
Un virus può essere dotato di un genoma a DNA o uno a RNA e pertanto vengono denominati rispettivamente virus a DNA o virus a RNA; la stragrande maggioranza sono a RNA. I virus delle piante tendono ad avere genomi composti da un singolo filamento di RNA mentre i batteriofagi spesso hanno un genoma a DNA a doppia elica.[87]
I genomi virali possono essere circolari, come nel poliomavirus, o lineari come negli adenovirus. Il tipo di acido nucleico è irrilevante per la forma del genoma. Tra virus a RNA e certi virus a DNA, il genoma è spesso diviso in parti distinte, nel qual caso esso è chiamato "segmentato". Per i virus a RNA, ogni segmento spesso codifica per una sola proteina e si trovano solitamente in un unico capside. Tuttavia, non è necessario che tutti i segmenti siano nello stesso virione perché il virus sia infettivo, come dimostrato dal virus del mosaico del tabacco e da molti altri virus delle piante.[67]
Un genoma virale, indipendentemente dal tipo di acido nucleico, è a singolo filamento oppure a doppio filamento. Genomi a singolo filamento consistono di un acido nucleico spaiato. Genomi a doppio filamento sono costituiti da due acidi nucleici complementari accoppiati. Quasi tutti i virus hanno un solo tipo di filamento, singolo oppure doppio, tuttavia alcune famiglie di virus, come quelli appartenenti agli Hepadnaviridae, contengono entrambi i tipi di filamento: ovvero hanno un genoma che è parzialmente a singolo filamento e parzialmente a doppio filamento.[87]
Per la maggior parte dei virus con genoma a RNA e alcuni con genomi a DNA a singolo filamento, i singoli filamenti sono classificati a senso-positivo (chiamato filamento positivo) o senso-negativo (chiamato filamento negativo), a seconda se sono complementari all'RNA messaggero virale (mRNA). L'RNA virale a senso positivo è nello stesso senso dell'mRNA virale e quindi almeno una parte di esso può essere tradotto immediatamente dalla cellula ospite. L'RNA virale a senso negativo è complementare all'mRNA e quindi deve essere convertito in RNA a senso-positivo tramite una RNA polimerasi RNA-dipendente prima della traduzione. La nomenclatura dei virus DNA con un genoma a un solo senso ssDNA è simile alla nomenclatura RNA, in quanto il filamento stampo per l'mRNA virale è complementare a esso (-) e il filamento codificante è una copia (+).[67] Tuttavia, diversi tipi di virus ssDNA e ssRNA hanno genomi che sono ambisenso e che la trascrizione può verificarsi fuori da entrambi i filamenti in un doppio filamento replicativo intermedio. Gli esempi includono i geminivirus, che sono i virus ssDNA delle piante e molti arenavirus che sono i virus ssRNA degli animali.[88]
La dimensione del genoma varia notevolmente tra le specie. I genomi virali più piccoli, i circoviruses ssDNA della famiglia dei circoviridae, codificano solo per due proteine e hanno una dimensione del genoma di due sole kilobase;[89] il più grande dei pandoravirus possono avere dimensioni del genoma di circa due megabasi che codificano per circa 2500 proteine.[90]
In generale, i virus a RNA hanno dimensioni del genoma più piccoli rispetto ai virus a DNA a causa di un più alto tasso di errore durante la replica e hanno un limite di dimensione massima superiore.[42] Al di là di questo limite, gli errori nel genoma durante la replica rendono il virus inutile o non infettivo. Per compensare questo, i virus a RNA spesso hanno un genoma segmentato: il genoma risulta pertanto diviso in molecole più piccole, riducendo così la possibilità che un errore in un singolo componente inabiliti l'intero genoma. Al contrario, i virus a DNA, in genere, possiedono genomi più grandi a causa della alta fedeltà dei loro enzimi di replicazione.[91] I virus a DNA a singolo filamento sono un'eccezione a questa regola, tuttavia, i tassi di mutazione di questi genomi possono avvicinarsi al caso estremo dei virus ssRNA.[92]
Come una mutazione o un riordinamento, possono portare a ceppi nuovi e altamente patogeni di influenza umana
I virus subiscono mutazioni genetiche attraverso diversi meccanismi. Tra questi, un processo chiamato deriva antigenica le cui basi individuali nel DNA o RNA mutano altre basi. La maggior parte di queste mutazioni puntiformi sono "silenziose", ovvero non cambiano la proteina che il gene codifica. Ma altri possono conferire vantaggi evolutivi quali la resistenza ai farmaci antivirali.[93][94] Lo spostamento antigenico si verifica quando vi è un cambiamento importante nel genoma del virus. Questo può essere il risultato di una ricombinazione o un riassortimento. Quando questo accade con i virus influenzali, possono verificarsi le pandemie.[95] I virus a RNA spesso esistono come quasispecie o sciami di virus della stessa specie, ma con una sequenza nucleotidica leggermente diversa del genoma. Tali quasispecie sono un obiettivo primario per la selezione naturale.[96]
Sequenze di genomi conferiscono vantaggi evolutivi; diversi ceppi di un virus con un genoma segmentato possono mischiare e combinare i geni e quindi produrre prole virus con caratteristiche uniche. Questo è chiamato riassortimento.[97]
La ricombinazione genetica è il processo mediante il quale un filamento di DNA viene rotto per poi essere unito al termine di una diversa molecola di DNA. Questo può verificarsi quando i virus infettano cellule simultaneamente e studi riguardanti l'evoluzione virale hanno dimostrato che la ricombinazione è stata molto frequente nelle specie studiate.[98] La ricombinazione è comune a entrambi i virus, sia a RNA sia a DNA.[99][100]
I virus non sono in grado di riprodursi attraverso la divisione cellulare poiché non sono cellule. Pertanto sfruttano il metabolismo e le risorse di una cellula ospite per produrre copie multiple di sé che si assemblano nella cellula.
Il ciclo di vita dei virus varia molto attraverso le diverse specie, ma vi sono sei fasi fondamentali:[101]
Un tipico ciclo di replicazione virale
Alcuni batteriofagi iniettano il loro genoma nelle cellule batteriche (non in scala)
L'attaccamento è un legame tra le proteine del capside virale e specifici recettori che si trovano sulla superficie cellulare della cellula ospite. La presenza di dati recettori sulla superficie determina la gamma degli ospiti di un virus. Ad esempio, l'HIV infetta una gamma limitata di leucociti umani. Questo perché le sue proteine di superficie, gp120, interagiscono specificamente con la molecola CD4, un recettore delle chemochine, che frequentemente si trova sulla superficie dei linfociti T. Questo meccanismo si è evoluto in modo da favorire quei virus che infettano solo le cellule in cui sono in grado di replicarsi. Il recettore attaccato può indurre la proteina dell'involucro virale a subire cambiamenti che si traducono nella fusione tra le membrane virali e cellulari, o cambiamenti alle proteine senza involucro di superficie del virus che non gli permettono di entrare.
L'ingresso segue l'attaccamento: virioni entrano nella cellula ospite mediante endocitosi mediata da recettori o attraverso la fusione delle membrane. Questo è spesso chiamato ingresso del virus. L'infezione delle cellule delle piante e dei funghi è diversa da quella delle cellule animali. Le piante hanno una parete cellulare rigida formata da cellulosa, mentre i funghi una di chitina, quindi la maggior parte dei virus possono entrare all'interno di queste cellule solo dopo aver perpetrato una rottura alla parete cellulare.[102] Tuttavia, quasi tutti i virus delle piante (come il virus mosaico del tabacco) possono anche passare direttamente da cellula a cellula, sotto forma di complessi a singolo filamento nucleoproteici, attraverso i pori chiamati plasmodesmi.[103] I batteri, come le piante, hanno pareti cellulari che un virus deve violare per infettare la cellula. Tuttavia, dato che le pareti delle cellule batteriche sono molto meno spesse delle pareti cellulari delle piante, per via delle loro dimensioni molto più piccole, alcuni virus possiedono evoluti meccanismi che iniettano il loro genoma nella cellula batterica attraverso la parete cellulare, mentre il capside virale rimane all'esterno.[104]
L'uncoating è un processo in cui il capside virale viene rimosso: questo può essere dovuto alla degradazione da parte degli enzimi virali o agli enzimi ospiti o per semplice dissociazione; il risultato finale è la liberazione dell'acido nucleico genomico virale.
La replicazione dei virus coinvolge principalmente la moltiplicazione del genoma. La replica consiste nella sintesi dell'RNA messaggero (mRNA) virale dai geni "early" (con eccezioni per i virus RNA a senso positivo), la sintesi proteica virale, il possibile montaggio delle proteine virali, quindi la replicazione del genoma virale. Questo può essere seguito, per i virus più complessi con genomi più grandi, da parte di uno o più cicli di sintesi di mRNA.
Assemblaggio, dopo l'auto-assemblaggio struttura-mediata delle particelle virali, alcune modifiche delle proteine si verifica spesso. Nei virus come l'HIV, questa modifica (a volte chiamato la maturazione) avviene dopo che il virus è stato rilasciato dalla cellula ospite.[105]
Rilascio, i virus possono essere rilasciati dalla cellula ospite per lisi, un processo che uccide la cellula rompendo la sua membrana e la parete cellulare, se presenti: questo avviene in molti batteri e di alcuni virus animali. Alcuni virus subiscono un ciclo lisogenico in cui il genoma virale è incorporato mediante la ricombinazione genetica in un luogo specifico dei cromosomi dell'ospite. Il genoma virale è quindi noto come "provirus" o, nel caso di un batteriofago "profago".[106] Ogni volta che l'ospite si divide, anche il genoma virale viene replicato. Il genoma virale è per lo più silente all'interno dell'ospite. Tuttavia, a un certo punto, il provirus o profago possono dar luogo al virus attivo, che può lisare le cellule ospiti.[107] I virus capsulati (ad esempio, l'HIV) di solito vengono rilasciati dalla cellula ospite per gemmazione. Durante questo processo il virus acquisisce il suo involucro, che è una parte modificata del plasma dell'ospite o della membrana interna.[108]
Il materiale genetico all'interno delle particelle del virus e il metodo con cui il materiale viene replicato, varia notevolmente tra i diversi tipi di virus.
La replicazione del genoma della maggior parte dei virus a DNA avviene nel nucleo della cellula. Se la cellula possiede il recettore appropriato sulla sua superficie, questi virus entrano, a volte, nella cellula per fusione diretta con la membrana cellulare (per esempio, l'herpes virus) o, più spesso. per endocitosi mediata da recettori. La maggior parte dei virus a DNA dipendono interamente dal DNA della cellula ospite e dai suoi strumenti di sintesi e lavorazione dell'RNA. Tuttavia, virus dotati di genomi più grandi, possono codificare gran parte di questi stessi strumenti. Negli eucarioti, il genoma virale deve attraversare la membrana nucleare della cellula per accedere ai meccanismi di replicazione, mentre nei batteri deve solo entrare nella cellula.[109]
La replicazione di solito avviene nel citoplasma. I virus a RNA possono essere classificati in quattro gruppi differenti in base alle modalità di replicazione. La polarità (se può o non può essere utilizzata direttamente dai ribosomi per produrre proteine) dei virus a singolo filamento di RNA determina in gran parte il meccanismo di replicazione; l'altro criterio principale è se il materiale genetico è a singolo o doppio filamento. Tutti i virus a RNA utilizzano i propri enzimi RNA replicasi per creare copie dei loro genomi.[110]
I virus a trascrizione inversa, possiedono ssRNA (Retroviridae, Metaviridae, Pseudoviridae) o dsDNA (Caulimoviridae, Hepadnaviridae) nelle loro particelle. Quelli con genomi a RNA (retrovirus), utilizzano un intermedio del DNA per replicare, mentre quelli con genomi di DNA (pararetroviruses) utilizzano un intermedio di RNA durante la replicazione del genoma. Entrambi i tipi utilizzano una trascrittasi inversa, o un enzima DNA polimerasi (RNA-dipendente) per effettuare la conversione dell'acido nucleico. I retrovirus integrano il DNA prodotto dalla trascrizione inversa nel genoma dell'ospite come provirus come parte del processo di replica; tuttavia, i pararetroviruses non lo fanno, anche se copie del genoma di particolari pararetroviruses infiltrati nei vegetali possono dar luogo a virus infettivo.[111] Essi sono suscettibili di farmaci antivirali che inibiscono l'enzima della trascrittasi inversa, ad esempio, la zidovudina e la lamivudina. Un esempio del primo tipo è l'HIV, che è un retrovirus. Esempi del secondo tipo sono il l'hepadnaviridae, che comprende il virus dell'epatite B.[112]
Vi è un'ampia serie di effetti strutturali e biochimici che i virus producono sulla cellula ospite.[113] Questi sono chiamati effetti citopatici.[114] La maggior parte delle infezioni da virus alla fine provocano la morte della cellula ospite. Le cause di morte sono la lisi cellulare, le alterazioni della membrana superficie della cellula e l'apoptosi.[115] Spesso la morte delle cellule è causata dalla cessazione delle sue normali attività a causa della soppressione da parte delle proteine specifiche del virus, che non tutti sono componenti della particella virale.[116]
Alcuni virus non provocano cambiamenti apparenti alla cellula infettata. Le cellule in cui il virus è latente e inattivo mostrano pochi segni di infezione e spesso vivono normalmente.[117] Ciò provoca infezioni persistenti e il virus rimane spesso in sospeso per molti mesi o anni. Ciò avviene frequentemente, ad esempio, con l'herpes virus.[118][119] Alcuni virus, come il virus di Epstein-Barr, possono indurre le cellule a proliferare senza provocare malignità,[120] mentre altri, come il papillomavirus, sono cause ormai stabilite di insorgenza di tumori.[121]
I virus sono di gran lunga le più abbondanti entità biologiche sulla Terra e superano tutte le altre messe insieme.[122] Essi sono in grado di infettare tutti i tipi di vita cellulare, tra cui animali, piante, batteri e funghi.[5] Tuttavia, le diverse tipologie di virus possono infettare solo una gamma limitata di ospiti e molti sono specie-specifici. Alcuni, come il virus del vaiolo umano, ad esempio, possono infettare solo una specie, in questo caso l'uomo,[123] e pertanto si dice che hanno una gamma di ospiti potenziali ristretta. Altri virus, come il virus della rabbia, possono infettare diverse specie di mammiferi e si dice che hanno una gamma di ospiti vasta.[124] I virus che infettano le piante sono innocui per gli animali e la maggior parte dei virus che infettano gli altri animali sono innocui per gli esseri umani.[125] La gamma dei possibili ospiti di alcuni batteriofagi è limitata a un unico ceppo di batteri e possono essere usati per rintracciare la fonte di focolai di infezioni, tramite un metodo chiamato fagotipizzazione.[126]
La classificazione cerca di descrivere la diversità dei virus nominandoli e raggruppandoli sulla base di somiglianze. Nel 1962, André Lwoff, Robert W. Horne e Paul Tournier per primi svilupparono un sistema di classificazione dei virus, in base al sistema gerarchico di Linneo.[127] Questo approccio si basa sul phylum, sulla classe, sull'ordine, sulla famiglia, sul genere e sulla specie. I virus sono stati raggruppati in base alle loro proprietà condivise (non quelle dei loro ospiti) e al tipo di acido nucleico che forma i loro genomi.[128] In seguito l'International Committee on Taxonomy of Viruses è stato costituito.
L'International Committee on Taxonomy of Viruses (ICTV) ha sviluppato l'attuale sistema di classificazione e ha formulato le linee guida che attribuiscono un peso maggiore ad alcune proprietà virali per mantenere l'uniformità della famiglia. È stata istituita un tassonomia unificata (un sistema universale di classificazione per i virus). Il rapporto lCTV 9[129] definisce il concetto di specie virale come il gruppo più basso nella gerarchia della ramificazione dei virus.[130] Tuttavia solo una piccola parte dei virus è stata studiata, con analisi di campioni provenienti da esseri umani che hanno dimostrato che il 20% circa delle sequenze virali recuperate non sono mai state viste prima, mentre campioni provenienti dall'ambiente, come dall'acqua di mare e sedimenti oceanici hanno rivelato la presenza di una grande maggioranza di sequenze totalmente nuove.[131]
Nell'attuale (2019) tassonomia ICTV, sono stati istituiti quattro domini: Duplodnaviria, Monodnaviria, Riboviria e Varidnaviria, cui si aggiungono altri virus non assegnati a nessun dominio. Il comitato non è responsabile della nomenclatura e della classificazione dei virus sotto il rango di specie, cioè di sottospecie, varietà, isolati, genotipi e sierotipi. Le regole di nomenclatura e tassonomia dell'ICTV sono riportate nell'International Code of Virus Classification and Nomenclature (ICVCN). In totale vi sono 7 ordini, 103 famiglie, 22 sottofamiglie, 455 generi, circa 2 827 specie e oltre 4 000 tipi non ancora classificati.[129][132][133]
La classificazione dei virus di Baltimore è basato sul meccanismo di produzione di mRNA. I virus devono generare mRNA dai loro genomi per produrre proteine e replicarsi, ma diversi meccanismi sono utilizzati per raggiungere questo obiettivo in ogni famiglia di virus. I genomi virali possono essere a singolo filamento (ss) o doppio filamento (ds), RNA o DNA, e possono o non possono utilizzare la trascrittasi inversa (RT). Inoltre, i virus ssRNA possono essere sia senso (+) sia antisenso (-). Questa classificazione pone i virus in sette gruppi:
I virus hanno differenti meccanismi per cui causano una malattia in un organismo, i quali dipendono in larga misura dalla specie virale. Meccanismi a livello cellulare comprendono principalmente la lisi cellulare, che comporta la rottura della cellula e la sua conseguente morte. Negli organismi pluricellulari, se un numero abbastanza consistente di cellule muoiono, l'intero organismo incomincerà a subirne gli effetti. Sebbene i virus causino la perdita dell'omeostasi, con conseguente insorgenza di una malattia, possono esisterne di relativamente innocui all'interno di un organismo. Un esempio potrebbe includere la capacità del virus dell'herpes simplex, responsabile dell'herpes labiale, di rimanere in uno stato inattivo nel corpo umano, una condizione denominata di "latenza"[138] che è una caratteristica dei virus herpes, tra cui il virus di Epstein-Barr, che provoca la febbre ghiandolare, e del virus della varicella-zoster, causa della varicella e dell'herpes zoster. La maggior parte delle persone al mondo, nella loro vita, sono state infettate con almeno uno di questi tipi di herpes virus.[139] Tuttavia, questi virus latenti possono talvolta essere utili, infatti la presenza del virus può aumentare l'immunità contro alcuni batteri patogeni, come lo Yersinia pestis.[140]
Alcuni virus possono causare un'infezione permanente o cronica, in cui i virus continuano a replicarsi nel corpo, nonostante i meccanismi di difesa dell'ospite.[141] Questo è comune nei virus dell'epatite B e nelle infezioni da virus dell'epatite C. Le persone con una infezione cronica sono note come vettori, in quanto fungono da serbatoi del virus infettivo.[142] Nelle popolazioni con un'alta percentuale di portatori, la malattia si dice che sia endemica.[143]
L'epidemiologia virale è una branca della scienza medica che si occupa di studiare la trasmissione e controllare le infezioni da virus negli esseri umani. La trasmissione di virus può essere verticale, cioè da madre a figlio, o orizzontale, cioè da persona a persona. Esempi di trasmissione verticale comprendono il virus dell'epatite B e l'HIV, in cui il bambino nasce già infettato con il virus.[144] Un altro, più raro, esempio è il virus della varicella zoster, che, pur causando infezioni relativamente lievi negli esseri umani, può essere fatale per il feto e il neonato.[145]
La trasmissione orizzontale è il meccanismo più comune di diffusione del virus nelle popolazioni. La trasmissione può verificarsi quando vi è scambio di fluidi corporei durante un rapporto sessuale, come nel caso dell'HIV; o quando vi è contaminazione di sangue infetto come durante una trasfusione o con lo scambio di aghi, come spesso si riscontra nella trasmissione dell'epatite C; lo scambio di saliva dalla bocca, per esempio può portare alla trasmissione del virus di Epstein-Barr; il norovirus può trasmettersi con l'ingestione di cibo o acqua contaminati; il virus dell'influenza per inalazione; mentre insetti vettori, come le zanzare, trasmettono all'organismo ospite alcuni virus come quello della febbre dengue. Il tasso o la velocità di trasmissione delle infezioni virali dipende da vari fattori che includono la densità della popolazione, il numero dei soggetti suscettibili (cioè quelli che non immuni),[146] la qualità dell'assistenza sanitaria e il tempo.[147] Quando un focolaio d'infezione provoca una percentuale insolitamente alta di casi in una popolazione o in una regione si parla di epidemia. Se i focolai sono diffusamente distribuiti nel pianeta si parla di pandemia.[148]
Tramite l'epidemiologia si tenta di interrompere la catena delle infezioni durante le epidemie di malattie virali.[149] Le misure di controllo utilizzate si basano sulla conoscenza delle modalità di trasmissione del virus. È fondamentale trovare la fonte o le fonti del focolaio in modo da identificare il virus, la cui catena di trasmissione può talvolta essere interrotta grazie a vaccini. Quando questi non sono disponibili, misure di igiene e di disinfezione possono essere rimedi efficaci. Le persone infette possono essere isolate dal resto della comunità e quelle che sono state esposte al virus poste in quarantena.[150] Per controllare l'epidemia di afta epizootica del 2001 in Gran Bretagna sono stati abbattuti migliaia di bovini.[151] Le infezioni virali degli esseri umani e degli altri animali hanno periodi di incubazione[152] che variano da pochi giorni a settimane e sono noti per la maggior parte delle infezioni.[152] Con una piccola sovrapposizione, al termine del periodo di incubazione vi è il periodo di contagiosità, in cui un individuo infetto può trasmettere la malattia.[152] Anche questo periodo è noto per molte infezioni virali e la conoscenza della durata di entrambi i periodi è importante nel controllo dei focolai.[153]
Le popolazioni dei nativi americani furono decimate dalle malattie contagiose, in particolare dal vaiolo, portate dai coloni europei in America. Non è chiaro quanti siano deceduti a causa delle malattie importate dagli stranieri dopo l'arrivo di Colombo, ma le stime indicano un possibile 70% della popolazione indigena.[senza fonte] Tutto ciò ha contribuito in modo significativo ai tentativi europei di spostare e sottomettere la popolazione nativa.[154]
Una pandemia è un'epidemia mondiale. La pandemia influenzale del 1918 (chiamata "influenza spagnola") che durò fino al 1919, fu catalogata come pandemia di livello 5, causata da un virus influenzale particolarmente aggressivo e mortale. Le vittime furono spesso giovani adulti sani, in contrasto con la maggior parte dei focolai di influenza che colpiscono prevalentemente i pazienti pediatrici, gli anziani o gli individui già indeboliti.[155] Le stime più datate indicano che questa pandemia abbia ucciso tra le 40 e le 50 milioni di persone,[156] mentre ricerche più recenti suggeriscono che tale dato potrebbe arrivare fino a 100 milioni di persone, il 5% della popolazione mondiale del 1918.[157]
La maggior parte dei ricercatori ritiene che l'origine del virus HIV sia nell'Africa sub-sahariana e che sia nato nel corso del XX secolo.[158] Anche la sua diffusione è oramai considerata una pandemia, con una stima di 38,6 milioni di persone infettate in tutto il mondo.[159] Le stime del Programma delle Nazioni Unite per l'AIDS/HIV (UNAIDS) e dell'Organizzazione Mondiale della Sanità (OMS) indicando che l'AIDS (la condizione dovuta al virus HIV) abbia ucciso più di 25 milioni di persone dal 5 giugno 1981, quando è stato riconosciuto, rendendola una delle epidemie più devastanti della storia.[160] Nel 2007 sono state stimate 2,7 milioni di nuove infezioni da HIV e 2 milioni di decessi correlati alla condizione.[161]
Diversi agenti patogeni virali altamente letali sono membri dei Filoviridae. I filovirus sono virus simili a filamenti che causano febbre emorragica, e comprendono l'Ebolavirus e il Marburgvirus. Il virus di Marburg, scoperto nel 1967, ha attirato l'attenzione dei media nel mese di aprile 2005 per un focolaio in Angola.[162] Il virus della malattia da virus Ebola ha causato epidemie intermittenti con alti tassi di mortalità, dal 1976 quando è stato identificato. Quella peggiore e più recente è stata l'epidemia di in Africa Occidentale del 2014.[163]
La prima linea di difesa dell'organismo contro i virus è il sistema immunitario innato. Questo comprende le cellule e gli altri meccanismi che difendono l'ospite da infezioni in modo non specifico. Ciò significa che le cellule del sistema innato riconoscono e rispondono agli agenti patogeni in modo generico ma, a differenza del sistema immunitario adattativo, esso non conferisce un'immunità duratura.[173]
L'interferenza dell'RNA è un importante difesa innata contro i virus.[174] Molti virus adottano una strategia di replica che coinvolge l'RNA a doppio filamento (dsRNA). Quando un virus infetta una cellula, rilascia la sua molecola di RNA che si lega immediatamente a un complesso proteico chiamato ribonucleasi III che taglia l'RNA in pezzi più piccoli. Un percorso biochimico, il complesso RISC, si attiva assicurando la sopravvivenza cellulare degradando l'mRNA virale. I rotavirus si sono evoluti per evitare questo meccanismo di difesa evitando di tagliare completamente l'RNA all'interno della cellula ma rilasciando una nuova produzione di mRNA attraverso i pori del capside interno della particella. In questo modo il loro dsRNA genomico resta protetto all'interno del nucleo del virione.[175][176]
Quando il sistema immunitario adattativo di un vertebrato incontra un virus, produce anticorpi specifici che si legano a esso e spesso sono in grado di renderlo non infettivo. Ciò è chiamata immunità umorale. Due tipi di anticorpi sono importanti. I primi, chiamati IgM, sono altamente efficaci nel neutralizzare i virus, ma vengono prodotti dalle cellule del sistema immunitario solo per poche settimane. I secondi, chiamati IgG, vengono prodotti di continuo. La presenza di IgM nel sangue dell'ospite viene utilizzato per diagnosticare la presenza di una infezione acuta, mentre le IgG indicano un'infezione passata.[177] Quando i test di immunità vengono effettuati, vengono misurati gli anticorpi IgG.[178]
Gli anticorpi possono continuare a essere un meccanismo di difesa efficace anche dopo che i virus sono riusciti a ottenere l'ingresso nella cellula ospite. Una proteina presente nelle cellule, chiamata TRIM21, è in grado di legarsi agli anticorpi sulla superficie della particella virale. Questo innesca la successiva distruzione del virus da parte degli enzimi del proteosoma della cellula.[179]
Una seconda difesa dei vertebrati contro i virus, si chiama immunità cellulo-mediata e coinvolge le cellule immunitarie conosciute come cellule T. Le cellule del corpo mostrano costantemente brevi frammenti delle loro proteine sulla superficie della cellula e se una cellula T riconosce un frammento virale sospetto, la cellula ospite viene distrutta dalle cellule T killer e le cellule T virus-specifiche proliferano. Le cellule, come i macrofagi sono specialisti in questa presentazione dell'antigene.[180] La produzione di interferone è anch'esso un importante meccanismo di difesa dell'ospite. Questo è un ormone prodotto dal corpo quando vi è la presenza di virus. Il suo ruolo nel sistema immunitario è complesso; ma sostanzialmente mette fine alla riproduzione del virus, uccidendo la cellula infetta e le sue vicine.[181]
Non tutte le infezioni da virus producono una risposta immunitaria protettiva in questo modo. L'HIV elude il sistema immunitario cambiando continuamente la sequenza aminoacidica delle proteine sulla superficie del virione. Questa tecnica è nota come "mutazione di fuga". Questi virus persistono nell'eludere il sistema immunitario, attraverso il sequestro e blocco della presentazione dell'antigene, la resistenza alle citochine, l'evasione delle attività delle cellule natural killer, evitare l'apoptosi e la sostituzione antigenica.[182] Altri virus, chiamati virus neurotropi, sfruttano la diffusione neurale in cui il sistema immunitario potrebbe non essere in grado di raggiungerli.
Poiché i virus utilizzano vie metaboliche vitali all'interno cellule ospiti per replicarsi, essi risultano difficili da eliminare senza usare farmaci che causino effetti tossici alle cellule dell'ospite. Gli approcci medici più efficaci per le malattie virali sono le vaccinazioni, in grado di fornire l'immunità alle infezioni, e i farmaci antivirali che interferiscono selettivamente con la replicazione del virus.
La vaccinazione è un modo economico ed efficace per prevenire le infezioni da virus. I vaccini sono stati usati per prevenire le infezioni virali molto prima della scoperta dei virus reali. Il loro utilizzo ha portato a un drastico calo della morbilità (l'ammalarsi) e della mortalità (morte) associate a infezioni virali come la poliomielite, il morbillo, la parotite e la rosolia.[183] Le infezioni di vaiolo sono state debellate.[184] A oggi, 2015, vi è la disponibilità di vaccini per oltre tredici infezioni virali che colpiscono gli esseri umani,[185] e molti altri vengono utilizzati per prevenire le infezioni virali degli animali.[186]
I vaccini possono essere costituiti da virus vivi, attenuati o morti, o da proteine virali (antigeni).[187] Le forme di vaccini vivi, contengono virus indeboliti che non causano la malattia ma, ciò nonostante, conferiscono l'immunità. I vaccini vivi possono essere pericolosi quando somministrati a persone con una debole immunità (individui immunocompromessi) poiché in queste persone il virus indebolito può provocare la malattia originale.[188] La biotecnologia e le tecniche di ingegneria genetica vengono utilizzate per la produzione di vaccini di subunità. Questi vaccini utilizzano solo le proteine del capside del virus. Il vaccino contro l'epatite B è un esempio di questo tipo di vaccino.[189] I vaccini di subunità sono sicuri per i pazienti immunocompromessi perché non possono in nessun modo provocare la malattia.[190] Il vaccino contro il virus della febbre gialla, un ceppo vivo attenuato e chiamato 17D, è probabilmente il più sicuro e il più efficace vaccino mai prodotto.[191]
I farmaci antivirali sono spesso analoghi nucleosidici (costruzioni di DNA falso), che i virus erroneamente incorporano nei loro genomi durante la replicazione. Il ciclo di vita del virus viene bloccata perché il DNA di nuova sintesi risulta inattivo. Infatti in questi analoghi mancano i gruppi ossidrilici, che, insieme con gli atomi di fosforo, si collegano insieme per formare la forte "spina dorsale" della molecola di DNA. Ciò prende il nome di interruzione della catena del DNA.[192] Esempi di analoghi nucleosidici sono l'aciclovir per il Virus dell'herpes simplex e la lamivudina per le infezioni da virus HIV e epatite B. L'aciclovir è uno dei più antichi farmaci antivirali e più frequentemente prescritti.[193] Altri farmaci antivirali utilizzano come bersaglio diverse fasi del ciclo di vita del virus. L'HIV dipende da un enzima proteolitico chiamato HIV-1 proteasi per farlo diventare completamente infettivo. Vi è un vasto gruppo di farmaci, chiamati inibitori della proteasi, che inattivano questo enzima.
L'epatite C è causata da un virus a RNA. Nell'80% delle persone infette la malattia risulta cronica e in assenza di trattamento restano infetti per tutta la loro vita. Tuttavia, vi è ora un trattamento efficace che utilizza un analogo nucleosidico, la ribavirina, combinato con l'interferone.[194] Il trattamento dei portatori cronici del virus dell'epatite B utilizzando una strategia simile utilizzando la lamivudina.[195]
I virus infettano tutta la vita cellulare anche se ogni specie ha una propria gamma di virus specifica in grado di infettarla.[196] Alcuni virus, chiamati satelliti, possono replicarsi solo all'interno delle cellule che sono già state infettate da un altro virus.[55]
I virus sono importanti a patogeni del bestiame. Malattie come l'afta epizootica e la febbre catarrale dei piccoli ruminanti sono causati da virus.[197] Gli animali da compagnia, come gatti, cani e cavalli, se non vaccinati, sono suscettibili di gravi infezioni virali. Il Parvovirus canina è causata da un piccolo virus a DNA e le infezioni sono spesso fatali nei cuccioli.[198] Come tutti gli invertebrati, le api sono suscettibili di molte infezioni virali.[199] Tuttavia, la maggior parte dei virus coesistono con il loro ospite e non causano segni o sintomi di malattia.[24]
Vi sono molti tipi di virus in grado di infettare il mondo vegetale, tuttavia il più delle volte causano solo una perdita di rendimento della produzione dei frutti e quindi non è economicamente vantaggioso cercare di controllarli. Virus si diffondono da pianta a pianta attraverso organismi, noti come vettori. Solitamente questi sono insetti, ma anche alcuni funghi, vermi nematodi e organismi unicellulari hanno dimostrato di poter essere vettori. Quando il controllo delle infezioni nelle coltivazioni è ritenuta economica, ad esempio per gli alberi da frutta, gli sforzi sono concentrati sull'uccidere i vettori e rimuovere ospiti alternativi, come le erbacce.[200] I virus delle piante non possono infettare gli esseri umani e altri animali poiché sono in grado di riprodursi solo all'interno di una cellula vegetale.[201]
I vegetali possiedono elaborati meccanismi di difesa contro i virus. Uno dei più efficaci è la presenza del gene cosiddetto di resistenza (R). Ogni gene R conferisce una resistenza a un particolare virus attivando aree localizzate di morte cellulare intorno alla cellula infetta, che spesso possono essere viste a occhio nudo come grandi macchie. ciò permette di interrompere la diffusione dell'infezione.[202] L'interferenza RNA è anch'essa un'efficace difesa delle piante.[203] Quando sono infette, le piante spesso producono disinfettanti naturali che uccidono i virus, come l'acido salicilico, l'ossido nitrico e specie reattive dell'ossigeno.[204]
Le particelle dei virus delle piante hanno importanti applicazioni sia nelle biotecnologie sia nelle nanotecnologie. Nella maggior parte dei virus delle piante, i capsidi appaiono come strutture semplici e robuste che possono essere prodotte in grandi quantità, sia per l'infezione di piante o mediante l'espressione in una varietà di sistemi eterologhi. Le particelle dei virus delle piante possono essere modificati geneticamente e chimicamente per incapsulare materiale estraneo e possono essere incorporati in strutture supramolecolari per l'uso nel campo delle biotecnologie.[205]
Micrografia a trasmissione elettronica di molteplici batteriofagi che attaccano una parete di un batterio
I batteriofagi (detti anche fagi) sono un gruppo comune e diversificato di virus e sono la forma più abbondante di entità biologica presente negli ambienti acquatici. Vi sono fino a dieci volte di questi virus negli oceani rispetto a quanti siano i batteri,[206] raggiungendo i valori di 250.000.000 batteriofagi per millilitro di acqua di mare.[207] Questi virus infettano batteri specifici legandosi ai recettori di superficie per poi entrare nella cellula. Nel giro di un breve lasso di tempo, in alcuni casi pochi minuti, la polimerasi batterica incomincia a tradurre l'mRNA virale in proteine. Queste proteine diventano sia nuovi virioni all'interno della cellula che aiutano il montaggio di nuovi virioni, sia proteine coinvolte nella lisi cellulare. Gli enzimi virali aiutano nella ripartizione della membrana cellulare, e, nel caso del fago T4, in poco più di venti minuti dopo l'infezione, oltre trecento fagi possono essere rilasciati.[208]
I principali batteri si difendono dai batteriofagi tramite la produzione di enzimi che distruggono il DNA estraneo. Questi enzimi, chiamati endonucleasi di restrizione, tagliano il DNA virale che batteriofagi iniettano nelle cellule batteriche.[209] I batteri contengono anche un sistema che utilizza sequenze CRISPR e che conserva frammenti del genoma dei virus che sono entrati in contatto con esso in passato; ciò gli consente di bloccare la replicazione del virus attraverso l'interferenza dell'RNA.[210][211] Questo sistema genetico fornisce ai batteri una immunità acquisita alle infezioni.
Alcuni virus si replicano all'interno archeobatteri: questi sono virus DNA a doppia elica con forme insolite e talvolta unici.[7][84] Sono stati studiati in maggior dettaglio negli archeobatteri termofili, in particolare gli ordini Sulfolobales e Thermoproteales.[212] Difese contro questi virus possono comportare l'interferenza dell'RNA da sequenze ripetute di DNA all'interno di genomi archei che riguardano i geni dei virus.[213][214]
Sebbene agli inizi i virus venivano studiati esclusivamente per i loro potenziali effetti patologici, oggi si usano approcci più ampi, avendo riconosciuto i ruoli importanti che rivestono nell'ambiente.
Un cucchiaino di acqua di mare contiene circa un milione di virus.[215] La maggior parte di questi sono batteriofagi, che sono innocui per le piante e gli animali ma sono in realtà essenziali per la regolazione degli ecosistemi d'acqua salata e di acqua dolce.[216] Essi infettano e distruggono i batteri delle comunità microbiche acquatiche e sono il più importante meccanismo di riciclaggio del carbonio nell'ambiente marino. Le molecole organiche rilasciate dalle cellule batteriche morte stimolano la crescita di nuovi batteri e alghe.[217] l'attività virale può anche contribuire alla pompa biologica, il processo attraverso il quale carbonio viene immagazzinato nelle profondità dell'oceano.[218]
I microrganismi costituiscono più del 90% della biomassa marina. Si stima che i virus uccidano circa il 20% di questa biomassa ogni giorno e che vi sia un numero 15 volte maggiore di virus negli oceani rispetto ai batteri e agli archeobatteri. I virus sono i principali agenti responsabili della rapida distruzione delle fioriture algali nocive,[219] che spesso uccidono altre forme di vita marina.[220] Il numero di virus negli oceani diminuisce più ci si sposta verso il largo e più ci si immerge in profondità, dove si trovano meno organismi ospiti.[218]
Ricercatori indiani suggeriscono che l'acqua del fiume Gange può rappresentare un'utile terapia nei confronti del COVID-19.[222] Infatti questa è ricca di batteriofagi (si ritiene contienga circa 1 100 tipi di batteriofagi)[223] che sono presenti in essa per il graduale scioglimento del permafrost himalayano nel fiume.[224] Inoltre, l'acqua di questo fiume mostra un'elevata alcalinità e alcune delle sue proprietà autopurificanti contribuiscono alla crescita dei batteriofagi.[223]
Il riscaldamento climatico osservato in Siberia e nell'Artico e lo scongelamento del permafrost potrebbero comportare il rilascio di agenti patogeni intrappolati nel permafrost nell'atmosfera, rappresentando un possibile meccanismo di genesi di virus che in futuro potrebbero emergere come conseguenza dello sviluppo del riscaldamento globale del nostro pianeta nei decenni a venire.[225]
I virus sono un importante mezzo naturale per il trasferimento di geni tra le diverse specie, aumentando la diversità genetica e la deriva evolutiva.[13] Si ritiene che i virus abbiano avuto un ruolo centrale nella prima evoluzione, prima che vi fosse la diversificazione dei batteri, degli Archea e degli eucarioti, nel momento in cui vi era sulla Terra l'ultimo antenato comune universale.[226] I virus sono ancora una delle più grandi riserve di diversità genetica inesplorate sulla Terra.[218]
I virus risultano importanti per lo studio della biologia molecolare e cellulare, in quanto forniscono sistemi semplici che possono essere utilizzati per manipolare e analizzare le funzioni delle cellule.[227] Lo studio e l'uso di virus hanno fornito preziose informazioni riguardo aspetti della biologia cellulare.[228] Ad esempio, i virus sono stati utili per lo studio della genetica e hanno aiutato a comprendere i meccanismi di base della genetica molecolare, come la replicazione del DNA, la trascrizione, l'elaborazione dell'RNA, la traduzione, il trasporto di membrana e l'immunologia.
I genetisti spesso usano virus come vettori per introdurre geni in cellule che stanno studiando. Ciò è utile per far sì che la cellula produca una sostanza estranea o per studiare l'effetto dell'introduzione di un nuovo gene nel genoma. In maniera simile, la viroterapia utilizza i virus come vettori per il trattamento di varie malattie in quanto sono in grado di incidere direttamente sulle cellule sul DNA. Questa tecnica sembra molto promettente nel trattamento dei tumori e nella terapia genica. Scienziati europei e orientali utilizzarono la terapia mediante i fagi come alternativa agli antibiotici per qualche tempo e l'interesse per questo approccio è in aumento per via dell'elevata resistenza antibiotica riscontrata in alcuni batteri patogeni.[229] L'espressione di proteine eterologhe da virus è la base di numerosi processi di fabbricazione che vengono utilizzati per la produzione di varie proteine, quali vaccini e anticorpi. Processi industriali sono stati recentemente sviluppati utilizzando vettori virali e un certo numero di proteine farmaceutiche sono in fase di studio clinico e pre-clinico.[230]
La viroterapia consiste nell'uso di virus geneticamente modificati per il trattamento della malattie.[231] I virus vengono modificati dagli scienziati in modo da attaccare e riprodursi nelle cellule tumorali distruggendole, ma non infettando le cellule sane. Il Talimogene laherparepvec (T-VEC), per esempio, è un virus herpes simplex a cui un gene, utilizzato per replicarsi nelle cellule sane, è stato eliminato e sostituito con gene umano (GM-CSF) che stimola la risposta immunitaria. Quando questo virus infetta le cellule tumorali le distrugge e la presenza del gene GM-CSF attrae le cellule dendritiche dai tessuti circostanti del corpo. Le cellule dendritiche elaborano le cellule tumorali morte e i loro componenti vengono "segnalati" alle altre cellule del sistema immunitario.[232] Ci si aspetta che, una volta terminati con successo gli studi clinici, questo virus possa ottenere l'approvazione per il trattamento del melanoma, un tumore della pelle.[233] I virus che sono stati riprogrammati per uccidere le cellule tumorali vengono chiamati virus oncolitici.[234]
Le attuali ricerche nel campo delle nanotecnologie promettono di rendere molto versatile l'utilizzo di virus. Dal punto di vista di uno scienziato dei materiali, i virus possono essere considerati come nanoparticelle organiche. La loro superficie è in grado di trasportare strumenti specifici volti a superare le barriere delle loro cellule ospiti. La dimensione e la forma del virus, e il numero e la natura dei gruppi funzionali sulla loro superficie, è definita con precisione. Una particolare qualità dei virus è che possono essere personalizzati per evoluzione diretta. Le potenti tecniche sviluppate dalle scienze della vita stanno diventando la base di approcci ingegneristici verso i nanomateriali e l'apertura di una vasta gamma di applicazioni che vanno ben oltre la biologia e la medicina.[235]
Per via della loro dimensione, forma e struttura chimica ben definite, i virus sono stati utilizzati come modelli per l'organizzazione dei materiali su scala nanometrica. Esempi recenti includono il lavoro presso lo United States Naval Research Laboratory di Washington, con particelle del virus del mosaico del fagiolo dall'occhio (CPMV) utilizzate per amplificare i segnali dei sensori basati su microarray di DNA. In questa esempio, le particelle virali separano i coloranti fluorescenti utilizzati per la segnalazione per prevenire la formazione di dimeri non fluorescenti che agiscono come smorzatori.[236] Un altro esempio è l'uso di una basetta di CPMV come nanoscala per l'elettronica molecolare.[237]
Molti virus possono essere sintetizzati "da zero" e il primo virus sintetico è stato creato nel 2002.[238] In effetti non è il virus che viene sintetizzato, ma piuttosto il suo genoma DNA (in caso di un virus DNA) o una copia cDNA del suo genoma (in caso di virus a RNA). Per molte famiglie di virus sintetici a DNA o RNA (una volta enzimaticamente convertito indietro dal cDNA sintetico) sono infettivi quando vengono introdotti in una cellula, cioè contengono tutte le informazioni necessarie per produrre nuovi virus. Questa tecnologia viene ora utilizzata per studiare nuove strategie vaccinali.[239] Le intere sequenze del genoma di 3843 virus diversi sono accessibili al pubblico in una banca dati online gestita dal National Institutes of Health.[240]
La capacità dei virus di causare epidemie devastanti nelle società umane ha portato alla preoccupazione che essi possano essere utilizzati come armi per la guerra biologica. Ulteriore preoccupazione è stata sollevata dalla ricreazione con successo del pericoloso virus dell'influenza spagnola in un laboratorio.[241] Il virus del vaiolo ha decimato numerose popolazioni nel corso della storia, prima della sua eliminazione. Vi sono solo due centri al mondo autorizzati dall'OMS a mantenere le scorte di questo virus: l'Istituto VECTOR in Russia e il Centro per la prevenzione e il controllo delle malattie negli Stati Uniti.[242] La paura che esso possa essere utilizzato come arma non può essere del tutto infondata.[242] Poiché il vaccino antivaioloso ha dimostrato di avere talvolta gravi effetti collaterali, non è più somministrato di routine in tutto il mondo, cosicché gran parte della popolazione umana moderna non possiede quasi nessuna difesa contro il virus e sarebbe quindi vulnerabile di fronte a una sua ricomparsa.[242]
Biota / Vitae / Eobiontes[246][247] - fattori viventi (sistemi in uno stato energetico di disequilibrio stazionario in grado di dirigere una serie di reazioni chimiche[248])
^Koonin EV, Starokadomskyy P (October 2016). "Are viruses alive? The replicator paradigm sheds decisive light on an old but misguided question". Studies in History and Philosophy of Biological and Biomedical Sciences. 59: 125–34. doi:10.1016/j.shpsc.2016.02.016. PMC 5406846. PMID 26965225
^From Nobel Lectures, Physics 1981–1990, (1993) Editor-in-Charge Tore Frängsmyr, Editor Gösta Ekspång, World Scientific Publishing Co., Singapore.
In 1887, Buist visualised one of the largest, Vaccinia virus, by optical microscopy after staining it. Vaccinia was not known to be a virus at that time. (Buist J.B. Vaccinia and Variola: a study of their life history Churchill, London)
^ Yu. G. Kuznetsov, A. J. Malkin, R. W. Lucas, M. Plomp, A. McPherson, Imaging of viruses by atomic force microscopy, in J Gen Virol, vol. 82, n. 9, 2001, pp. 2025-2034, PMID11514711.
^(EN) Nadège Philippe et al., Pandoraviruses: Amoeba Viruses with Genomes Up to 2.5 Mb Reaching That of Parasitic Eukaryotes (abstract), in Science, vol. 341, n. 6143, American Association for the Advancement of Science, 19 luglio 2013, pp. 281-286, DOI:10.1126/science.1239181. URL consultato il 12 agosto 2018.
^ Hampson AW, Mackenzie JS, The influenza viruses, in Med. J. Aust., vol. 185, 10 Suppl, 2006, pp. S39–43, PMID17115950.
^ Metzner KJ, Detection and significance of minority quasispecies of drug-resistant HIV-1, in J HIV Ther, vol. 11, n. 4, 2006, pp. 74-81, PMID17578210.
^ab King AMQ, Lefkowitz E, Adams MJ, Carstens EB, Virus Taxonomy: Ninth Report of the International Committee on Taxonomy of Viruses, Elsevier, 2011, p. 6, ISBN0-12-384684-6.
As defined therein, "A species is the lowest taxonomic level in the hierarchy approved by the ICTV. A species is a monophyletic group of viruses whose properties can be distinguished from those of other species by multiple criteria."
This Excel file contains the official ICTV Master Species list for 2014 and lists all approved virus taxa. This is version 11 of the MSL published on 24 August 2014. Retrieved on 24 December 2014
^ Baltimore D, The strategy of RNA viruses, in Harvey Lect., 70 Series, 1974, pp. 57-74, PMID4377923.
^ Rodrigues C, Deshmukh M, Jacob T, Nukala R, Menon S, Mehta A, Significance of HBV DNA by PCR over serological markers of HBV in acute and chronic patients, in Indian journal of medical microbiology, vol. 19, n. 3, 2001, pp. 141-4, PMID17664817.
^ Sauerbrei A, Wutzler P, The congenital varicella syndrome, in Journal of perinatology : official journal of the California Perinatal Association, vol. 20, 8 Pt 1, 2000, pp. 548-54, PMID11190597.
^(RU) Platonov AE, (The influence of weather conditions on the epidemiology of vector-borne diseases by the example of West Nile fever in Russia), in Vestn. Akad. Med. Nauk SSSR, n. 2, 2006, pp. 25-9, PMID16544901.
^Status of the global HIV epidemic (PDF), su data.unaids.org, UNAIDS, 2008. URL consultato il 19 dicembre 2014 (archiviato dall'url originale il 22 novembre 2015).
^ Bellon M, Nicot C, Telomerase: a crucial player in HTLV-I-induced human T-cell leukemia, in Cancer genomics & proteomics, vol. 4, n. 1, 2007, pp. 21-5, PMID17726237.
^Hall, A. J., Jepson, P. D., Goodman, S. J. & Harkonen, T. "Phocine distemper virus in the North and European Seas — data and models, nature and nurture". Biol. Conserv. 131, 221–229 (2006)
^(EN) Harvey Lodish, Arnold Berk, S. Lawrence Zipursky, Paul Matsudaira, David Baltimore e James Darnell, Section 6.3 Viruses: Structure, Function, and Uses, in Molecular Cell Biology, 4ª ed., New York, W. H. Freeman, 2000. URL consultato il 12 agosto 2018.
^ Gleba, YY; Giritch, A, Plant Viral Vectors for Protein Expression, in Recent Advances in Plant Virology, Caister Academic Press, 2011, ISBN978-1-904455-75-2.
^ Luketa S., New views on the megaclassification of life (PDF), in Protistology, vol. 7, n. 4, 2012, pp. 218–237. URL consultato il 15 novembre 2022 (archiviato dall'url originale il 2 aprile 2015).
(EN) Leslie Collier, Albert Balows e Max Sussman, Virology, in Brian H Mahy e Leslie Collier (a cura di), Topley and Wilson's Microbiology and Microbial Infections, vol. 1, 9ª ed., Arnold, 1998, ISBN0-340-66316-2.
(EN) Knipe, David M; Howley, Peter M; Griffin, Diane E; Lamb, Robert A; Martin, Malcolm A; Roizman, Bernard; Straus Stephen E. (2007) Fields Virology, Lippincott Williams & Wilkins. ISBN 0-7817-6060-7.
(EN) S. Casjens, Assembly of Bacterial Viruses, in Brian W. J. Mahy e Marc H. V. Van Regenmortel (a cura di), Desk Encyclopedia of General Virology, Boston, Academic Press, 2010, pp. 167-174, ISBN0123751624.
(EN) F. Fenner, History of Virology: Vertebrate Viruses, in Brian W.J. Mahy e Marc H.V. Van Regenmortel (a cura di), Desk Encyclopedia of General Virology, 1ª ed., Oxford, UK, Academic Press, 2009, pp. 15-19, ISBN0-12-375146-2.
(EN) P. Forterre, Origin of Viruses, in Brian W.J. Mahy e Marc H.V. Van Regenmortel (a cura di), Desk Encyclopedia of General Virology, Oxford, Academic Press, 2009, pp. 23. ..., ISBN0-12-375146-2.
(EN) King AMQ, Lefkowitz E, Adams MJ, Carstens EB, Virus Taxonomy: Ninth Report of the International Committee on Taxonomy of Viruses, Elsevier, 2011, p. 6, ISBN0-12-384684-6.
(EN) Fisher, Bruce; Harvey, Richard P.; Champe, Pamela C., Lippincott's Illustrated Reviews: Microbiology (Lippincott's Illustrated Reviews Series), Hagerstwon, MD, Lippincott Williams & Wilkins, 2007, pp. 367–392, ISBN0-7817-8215-5.
(EN) Gleba, YY; Giritch, A, Plant Viral Vectors for Protein Expression, in Recent Advances in Plant Virology, Caister Academic Press, 2011, ISBN978-1-904455-75-2.
(EN) Witzany G (Ed), Viruses: Essential Agents of Life, Springer, 2012, ISBN978-94-007-4898-9.








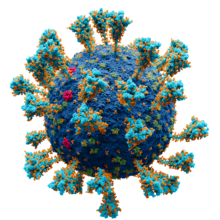












 Wikiquote contiene citazioni di o su virus
Wikiquote contiene citazioni di o su virus Wikizionario contiene il lemma di dizionario «virus»
Wikizionario contiene il lemma di dizionario «virus» Wikimedia Commons contiene immagini o altri file su virus
Wikimedia Commons contiene immagini o altri file su virus Wikispecies contiene informazioni su virus
Wikispecies contiene informazioni su virus